
It’s a precarious gamble in paleontology naming a new species of dinosaur from a single, isolated tooth. Things get pretty confusing after a while, which is why modern paleontologists don’t really do this anymore, and use unique, diagnostic features in the bones plus the teeth of fossil animals as a basis for describing a new species. A good example of this can be seen in the story of Troodon, a dinosaur many people think they know… But do we really?
The name Troodon formosus was given to a tooth collected in Montana in 1855. It was named by the forefather of American paleontology Joseph Leidy the following year, making it one of the first dinosaur remains known from North America (Leidy, 1856). It’s hard to blame Leidy for naming a whole new animal from a single tooth, since the science of paleontology was still in its infancy, and nothing like this tooth had ever been seen before. How was he supposed to know there was any other kind of animal out there that had teeth similar to this? The Troodon tooth came from the Judith River Formation, making it between 77.5 and 76.5 million years old or so, and went through a decades-long identity crisis which, in a way, continues to this day. Troodon was first identified as a prehistoric lizard, then a type of pachycephalosaur, but these ideas were overturned in the 30’s and 40’s when the first troodontid skeletal material with associated teeth was discovered in Alberta’s Dinosaur Park Formation, showing that Troodon was actually a small theropod. The legendary Charlie Sternberg named the Alberta material Stenonychosaurus inequalis, a roughly dog-sized animal with hollow, birdlike bones and teeth essentially the same as those from the Montanan Troodon (Sternberg, 1932).

The cutting edge of a troodontid tooth fragment from northwestern Alberta. Note the prominent, hooked denticles. Photo by Nicholas Carter
Throughout the late 20th century, more and more Stenonychosaurus remains were being discovered in Alberta, which included a few rare skull fragments and braincases. Additionally, Troodon-like teeth were found in end-Cretaceous formations in the United States, and were given the name Pectinodon bakkeri (Carpenter, 1982) The skeletal fossils clued paleontologists into the fact that troodontids had big eyes, a big brain, and were altogether very birdlike. Paleontologist Dale Russell famously brought Stenonychosaurus to the world in a description accompanied by a featherless sculpture of the dinosaur (Russell & Séguin 1982). Russell apparently saw something a little humanlike in the big brain and flexible first digit of this dinosaur, and commissioned the sculpture of a hypothetical post-Cretaceous troodontid that, having never been struck down by the meteor, went on to survive and develop its perceived humanlike traits even further. This theoretical and somewhat nightmarish beast, dubbed the “dinosauroid”, defies easy description.
The following decade, Phil Currie not only recognized the birdlike features of troodontids, but also couldn’t find anything significant that separated the different troodontids of North America. So, he lumped Stenonychosaurus and Pectinodon under the name Troodon formosus (Currie, 1987). This, however, was not to last. Paleontologists, including Currie himself, figured it was safer to keep the animals that were once the Albertan Stenonychosaurus and end-Cretaceous Pectinodon as their own species within the Troodon genus, and so they became Troodon inequalis and “Troodon bakkeri” respectively (Currie, 2005). This uncertainty over what is and isn’t Troodon formosus stems from the fact that there’s only the original Judith River tooth, and there may not be much about that tooth distinguishes the owner of it from its troodontid relatives. This uncertainty has even led some paleontologists to conclude that, since there might not be anything diagnostic about the original Troodon specimen, perhaps we shouldn’t really be using that name till things are finally sorted out.
There’s still work to be done to get the clearest possible picture on the diversity of North American troodontids, but we’re getting there bit by bit. In a 2017 study, Aaron van der Reest and Phil Currie argued that the Dinosaur Park Formation troodontid remains should not be referred to as Troodon, and not only resurrected the taxon Stenonychosaurus, but split some fossils from higher-up in this formation into a new genus called Latenivenatrix mcmasterae (van der Reest & Currie, 2017).
Troodontid remains from Alberta’s Horseshoe Canyon Formation, both younger and slightly different in anatomy than the Dinosaur Park fossils, were finally described by our assistant curator Derek Larson and colleagues in 2017 and given the name Albertavenator curriei -the specific name being after Dr. Currie, unsurprisingly a big troodontid fan (Evans et al., 2017). A few other species have been described from American remains, and large troodontid fossils of an undetermined species have even been found in northern Alaska (Fiorillo, 2008). Closer to us at the Currie Museum, troodontid teeth have been found in the Wapiti Formation of northwestern Alberta, but sadly skeletal remains still elude us- for now (Fanti & Miyashita, 2009). So who knows what species we have up here? It’s also worth mentioning that a generous wealth of troodontid species, some preserved in amazing detail, have been recovered from Asia ever since the 1920’s.
So what do we know about the evolution and biology of this group of dinosaurs? The troodontids were fairly similar in general form to the dromaeosaurs (or ‘raptors’, though some ornithologists don’t appreciate having the name stolen), being small to medium theropods with long grasping fingers, toothy snouts, big brains, and an enlarged ‘killing claw’ that was generally held aloft. Troodontids, though, tended to more slender in build but generally larger than the dromaeosaurs they shared their habitats with. They downsized the killing claw by comparison, but invested a lot in the eyes and brains department. This made their skulls, as well as the rest of the body inside and out, very birdlike. In fact, troodontids, dromaeosaurs, and birds all form an evolutionary group together called the Paraves, meaning they’re all, more or less, each other’s closest relatives.
The brains of troodontids have long been one of the major points of interest when this family of dinosaurs is discussed, with some of the highest brain-to-body size ratios of any non-avian dinosaur group. Does this mean that they rivalled mammals, even primates, in intelligence? Was Russell’s terrifying ‘dinosauroid’ really not so far off the mark? Well, as we talked about in the Velociraptor article, big-brained dinosaurs almost certainly didn’t reach the heights of modern mammals, corvids, or parrots in their cognition. Think more chicken, ostrich, or opossum-levels of intelligence, which is still pretty good considering the more ‘reptilian’ mental capabilities of most other dinosaurs. But I think it’s unfair to judge dinosaurs by our narrow standards
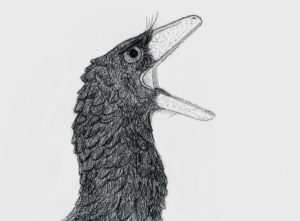
To the casual observer transported back to late Cretaceous Alberta, troodontids would have looked essentially like big, snappy birds. By Nicholas Carter
– they were extremely good at doing what they did for far longer than humans have been around.
Since the size of troodontid species ranged from about the size of a duck to the size of a deer, and the ecosystems they inhabited included arid deserts, swampy bayous, and temperate woodlands, there was probably a fair amount of variation in their lifestyles. Some may have had to become more specialized in their diet and behaviour based on what resources were available to them. One common feature throughout the group is fairly large sockets, and while this doesn’t necessarily mean that troodontids had big eyeballs to match, they certainly could have. For decades it’s been suggested that troodontids with large orbits may have been nocturnal, using their big eyes like a flightless owl to see in the low light as they hunted. This is bolstered by the fact that troodontids are some of the most common dinosaurs known from Arctic regions like Alaska. Since these small theropods were likely incapable of migrating out of this region during the winter like the larger hadrosaurs and horned dinosaur might have, they simply would’ve had to stay and make due (Bell & Snively, 2008). While the Arctic Circle may not have been as cold back then as it is today, it still had the 24-hour periods of darkness for large chunks of the year, so perhaps the big-eyed troodontids thrived in the endless winter night there millions of years ago. However, other species further south may have been less specialized in their hunting hours, and could have had big eyes just to see their prey in any lighting situation, much like diurnal owls and raptors today.

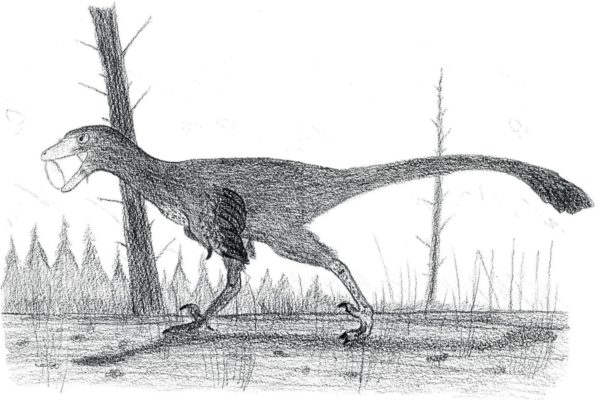
A northern troodontid hunkers down amidst a winter snowfall. We don’t know if northern species may have had lighter plumage than their southern cousins, but it’s tempting to imagine they did. By Nicholas Carter
There’s also an age-old debate as to what kind of prey items troodontids were taking advantage of, ever since that first tooth was looked upon by Joseph Leidy so long ago. While sharp like a typical carnivorous animal tooth, the teeth of bigger troodontids usually have very pronounced, slightly hooked denticles along the cutting edges. Some paleontologists saw more of a similarity in troodontid teeth to those of herbivorous reptiles such as iguana lizards and dome-headed dinosaurs, whose teeth are less pointed but also bear pretty big serrations. So, the idea the troodontids may have been omnivores, or perhaps even herbivores, has been tossed around over the years (Holtz et al., 1998). This was supported by the fact that troodontids appear to have weaker teeth compared to dromaeosaurs of similar size. However, with their raptorial claws, highly developed senses, and teeth still capable of processing flesh, the idea of troodontids as plant-eaters has mostly been discarded.
So what kind of prey did they tackle, then? Well, a good way to think of it is to look at what we know was available for eating at the time, and ask ourselves what troodontids were definitely not capable of eating, and everything else is theoretically fair game. The Cretaceous world was teeming with all sorts of small, easily processed food items, so they probably had lots of options. We know that troodontids probably didn’t hunt in the same manner as their dromaeosaur cousins, who with their strong jaws and dagger-like teeth could confidently bite into and tear a chunk from whatever sort of prey item they fancied, whether it was alive and kicking or not. Since troodontids don’t seem have been able to stand up to violently struggling prey, and their teeth weren’t so great for biting into flesh and bone with heedless abandon, it seems like they probably would have targeted smaller creatures that could be easily take down without much fuss (Torices et al., 2018). This may have included small mammals and birds, fish, lizards and snakes, and perhaps even invertebrates. It’s also of interest that troodontid teeth often appear in places that are thought to have been nesting sites for large herbivorous dinosaurs, such as Alberta’s own Devil’s Coulee or Kleskun Hills (Fanti & Miyashita, 2009). It could have been that these small predators were skulking around the region during nesting season, and would sneak in and nab an egg or newborn from the nest when they could.
A troodontid sneakily makes off with an unhatched dinosaur egg. By Nicolas Carter, based on Scott Hartman’s skeletal
Troodontids are still of great interest to paleontologists today. These dinosaurs have played a major part in understanding the evolution of birds, and early forms from China, wonderfully preserved with feathers intact, keep on providing more and more knowledge on the origins of birdlike traits in dinosaurs. Species from North America continue to prove enigmatic in their anatomy, lifestyle, and classification- ironic for a family of dinosaurs known of for so long. Time and effort by paleontologists with surely give us more insights into these fascinating dinosaurs. That’s what’s going on with Troodon.
By Nicholas Carter
References
Bell PR, Snively E. 2008. Polar dinosaurs on parade: a review of dinosaur migration. Alcheringa 32 (3):271-284
Carpenter, K. (1982). “Baby dinosaurs from the Late Cretaceous Lance and Hell Creek formations and a description of a new species of theropod” (PDF). Contributions to Geology, University of Wyoming. 20 (2): 123–134
Currie, P. J. (1987). “Bird-like characteristics of the jaws and teeth of troodontid theropods (Dinosauria, Saurischia)”. Journal of Vertebrate Paleontology. 7: 72–81
Currie, P. (2005). “Theropods, including birds.” in Currie and Koppelhus (eds). Dinosaur Provincial Park, a spectacular ecosystem revealed, Part Two, Flora and Fauna from the park.Indiana University Press, Bloomington. Pp 367–397
Evans, D. C.; Cullen, T.M.; Larson, D.W.; Rego, A. (2017). “A new species of troodontid theropod (Dinosauria: Maniraptora) from the Horseshoe Canyon Formation (Maastrichtian) of Alberta, Canada”. Canadian Journal of Earth Sciences. 54 (8): 813–826
Fanti, F. and Miyashita, T. 2009. A high latitude vertebrate fossil assemblage from the Late Cretaceous of west-central Alberta, Canada: evidence for dinosaur nesting and vertebrate latitudinal gradient. Palaeogeography, Palaeoclimatology, Palaeoecology. 275:37-53
Fiorillo, Anthony R. (2008) “On the Occurrence of Exceptionally Large Teeth of Troodon (Dinosauria: Saurischia) from the Late Cretaceous of Northern Alaska” Palaios volume 23 pp.322-328
Holtz, Thomas R., Brinkman, Daniel L., Chandler, Chistine L. (1998) Denticle Morphometrics and a Possibly Omnivorous Feeding Habit for the Theropod Dinosaur Troodon. Gaia number 15. December 1998. pp. 159–166
Leidy, J. 1856. Notices of remains of extinct reptile and fishes, discovered by Dr. F. V. Hayden in the bad lands of the Judith River, Nebraska Territory. Proceedings of the Academy of Natural Sciences, 8: 72-73
Russell, D. A.; Séguin, R. (1982). “Reconstruction of the small Cretaceous theropod Stenonychosaurus inequalis and a hypothetical dinosauroid”. Syllogeus. 37: 1–43
Sternberg, C.M. 1932. Two new theropod dinosaurs from the Belly River Formation of Alberta. Canadian Field-Naturalist, 46: 99-105
Torices, Angelica; Wilkinson, Ryan; Arbour, Victoria M; Ruiz-Omeñaca, Jose Ignacio; Currie, Philip J. (2018) Puncture-and-Pull Biomechanics in the Teeth of Predatory Coelurosaurian Dinosaurs. Current Biology; DOI: 10.1016/j.cub.2018.03.042
van der Reest, A. J.; Currie, P. J. (2017). “Troodontids (Theropoda) from the Dinosaur Park Formation, Alberta, with a description of a unique new taxon: implications for deinonychosaur diversity in North America”. Canadian Journal of Earth Sciences. 54 (9): 919–935