Let’s face it, we might not ever fully and completely know what every species of non-avian dinosaur looked like when it was alive. Heck, we probably won’t even discover every dinosaur that ever lived. Fossilizing is all about dying in the right place at the right time, and entire species probably came and went without leaving any trace of a geological record. It might seem sad that some dinosaurs lived and died with nothing to remember them by, but is recognition by humans really the only honor that a species gets in the course of time? Well, that’s a topic for another time, so let’s go back to the life appearance of dinosaurs.
We know a surprising amount about what dinosaurs looked like when they were alive considering we mostly just have their bones. But that’s not to say that’s all we have, and paleontologists can infer a lot from studying fossil skeletons and comparing them to modern day animals. The science of comparative anatomy is one of the foundations of paleontology, and is how the earliest paleontologists were able to figure out the basics of what extinct animals looked like. All vertebrate animals generally follow the same skeletal blueprint with just variations on the ancestral theme, so knowing the anatomy of living animals provides hints at what extinct ones looked like.
Other things like preserved soft-tissue impressions really help to literally ‘flesh out’ dinosaurs in our minds. Soft-tissue traces aren’t as uncommon as you might think, and we have preserved impressions of skin, feathers, and other bits from a variety of dinosaur taxa. One thing we must remember, though, is that the skin impressions that sometimes accompany dinosaur skeletons are not necessarily how they would appear when the animal was alive. Desiccation, unnatural posturing, decomposition, and taphonomic forces can potentially give us a distorted view of a dinosaur’s body covering.
So, from all of this, what do we really know about the life appearance of dinosaurs, and how does it differ from the conventional pop culture myths? Well, it’s been an evolving process throughout the decades. What the Victorians thought they knew about the appearance of dinosaurs is, in many ways, very different from what we think we know. We’ve (hopefully) moved closer to the facts over that time.
One of the big changes in dinosaur depictions since then has been our knowledge of their posture. Up until the 1970’s, it was standard fare for quadrupedal dinosaurs to be shown in a semi-sprawled posture with their tails dragging on the ground, and for bipeds to exhibit that upright kangaroo-style pose with the tail as a sort of third leg. The quadruped pose was mostly based on observations of modern four-legged reptiles like lizards and crocodiles. It was though that, since dinosaurs were reptiles too, they must have held their bodies in similar ways. The bipedal Godzilla-pose was inspired by kangaroos, since early paleontologists couldn’t think of any other way that a large animal with hind legs longer than its front legs would stand and move. An unbiased look at the way dinosaur bones would have fit together in life, combined with evidence from trackways (prints are generally close together and tail-drags are nearly always absent) and observing how modern avian dinosaurs walk has changed things. Dinosaurs normally carried their backbones and hips parallel to the ground, with tails held out for balance and heads in an upright, alert pose. Could many species get themselves into other postures from time to time when the need or desire struck them? Absolutely. But the default stance for dinosaurs was not the sluggish, sprawled, giant lizard pose that thankfully is increasingly absent in books, TV, and movies.
On a related note, dinosaur hands and feet are so often depicted wrong in the media and toys, which often give large species either stumpy, elephant-like feet or primitive lizard feet. For such large animals, the walking parts of dinosaurs were actually sort of dainty. Dinosaurs had a digitigrade posture, meaning only their toes up to the joint with the metatarsals contacted the ground. The rest of the foot and ankle were held upright in the same manner as a dog or a bird (the latter of which don’t have ‘backwards knees’ as is often heard. What you’re seeing there is actually the ankle joint, and the knees of birds rarely show beyond the feathers or body wall).
Dinosaurs normally had anywhere from 3-4 claws on the feet, which tended to form broad ‘hoofs’ in big herbivores. In theropods the toe claw sheathes were likely not too sharp, probably resembling that of big ground-dwelling birds. One obvious exception is the raptorial claws of dromaeosaurs and troodontids, which were probably much like the talons of modern raptors. The hands of dinosaurs came in all shapes and sizes depending on their lineage and lifestyle, but in most groups the palms of the hands didn’t face ‘downwards’ like most people automatically do when pretending to be a dinosaur. Humans being mammals, we have the luxury of flexible lower arms that can turn a fair ways. Instead, the inner face of most dinosaur hands pointed inwards, and birdlike theropods could somewhat ‘fold’ their wrists like the wings of a bird. Many dinosaur lineages reduced the number of fingers over time down to only two or three, but in groups that still had four or five, only the inner three fingers would have borne claws. Look closely at the hands of crocodilians and you’ll see the same thing. Preserved trackways confirm that larger dinosaurs (rhino-sized things and upwards) had fleshy pads behind their toes that helped these large animals to cushion their weight. Elephants and rhinos today show similar adaptations. This also means that most dinosaurs probably didn’t make thunderous stomping noises as they walked.
Dinosaur tails sometimes get shown as skinny, rat-like appendages slithering and curling at random or, perhaps worse, as merely ‘dead weight’. In reality, the tails of dinosaurs served all kinds of purposes, including balance, locomotion, defense, and social signaling. Most species aside from birds had large muscles at the base of the tail that connected to the femur bones. This would have made the tail a thick, muscular structure something more like a crocodile’s tail minus the lateral compression for swimming (Persons and Currie, 2011). Advanced carnivores had somewhat interlocking tail vertebrae that would have reduced the flopping of the tail as they moved, and derived ornithischian groups independently evolved ossified tendons that ran along the back, hips, and tail, making it a fairly stiff structure.
On that note, dinosaurs in popular media are still falling victim to the act of shrink-wrapping. This is style of dinosaur reconstruction that arose during the 70’s and 80’s where rigorous attention to the detail of the skeleton was overly-reflected on the animal’s external anatomy, resulting in essentially a walking dinosaur skeleton covered in skin. Now, we might never know the full extent of dinosaur soft tissue, but inferences from living animals can tell us that many reconstructions need to be bulked up with muscle and fat in at least some areas. Herbivorous dinosaurs had cheeks analogous to those of mammals that helped prevent food from tumbling out the sides of the mouth (Nabavizadeh, 2019). Then there’s the whole can of worms that is dinosaur soft-tissue display features and such. Things like soft crests, wattles, dewlaps, flaps, ridges, frills, and such may or may not have been present on all sorts of species, and we have evidence of a few examples in some things like ‘mummified’ hadrosaurs (Osborn, 1909)(Osborn, 1912).
Thanks to the preserved remains of the bony ring that supported the eyeball, we know roughly what size many dinosaur eyes were, and in many species the eyeball was much smaller than the eye socket. Most dinosaurs, though, were likely pretty visual animals. We don’t know exactly what the eyeball looked like, but the sclera of the eye almost certainly would’ve been either black or colourful as it is in crocodiles and birds, as opposed to white as in mammals. Most non-mammalian tetrapods have a nictitating membrane (that transparent ‘third eyelid’ that closes across the eye), and dinosaurs probably did as well.
Dinosaur colour is another thing we’re getting better and better at figuring out. It was once thought that we could only guess at what colour dinosaurs were, and while this still mostly true, we can narrow down the possibilities more so these days. The study of feathers from small Chinese theropods has revealed fossilized melanosomes- the organelles that produce the dark pigment melanin. Some also hit at iridescent qualities in the structure of the feathers. A handful of beautifully preserved ornithischians have faint traces of melanosome-bearing material from the skin preserved too. While we aren’t getting the full picture of dinosaur colouration yet since other factors contribute to colour, we’re mostly seeing dinosaurs in brown, dull red, and tan with some speckling here and there. It seems like at least some herbivores were counter-shaded to help avoid the notice of predators, and some Paraves like Anchiornis had striking red and white highlights in their feathers (Li et al., 2010).
The presence of scales, feathers, and bare skin is a sensitive topic in paleontology right now, with scientists trying to work out when exactly feathers arose in the dinosaur ‘family tree’, and if they’d ever been secondarily lost. So far, impressions of both scales and feather-like ‘fuzz’ are known from most major dinosaur lineages except sauropods and armored dinosaurs, for which we have only scales. It’s a complicated problem, and we’ll save the evolution and diversity for feathers for another day. Impressions of dinosaur skin normally show small, round, non-overlapping scales shaped vaguely like tiny hexagons. Some groups like ceratopsids often had large ‘tubercles’ surrounded by a mosaic of smaller scales. Some hadrosaurs had sections of their scaly skin arranged into series of bumps or folds. Large plates, knobs, and spikes along the body are mostly seen in the armored thyreophorans, though some titanosaur sauropods had simple ones as well. Theropods also typically fit the small, smooth scale mold, aside from at least a few ceratosaurs like Carnotaurus and Ceratosaurus which had bony knobs along the body.
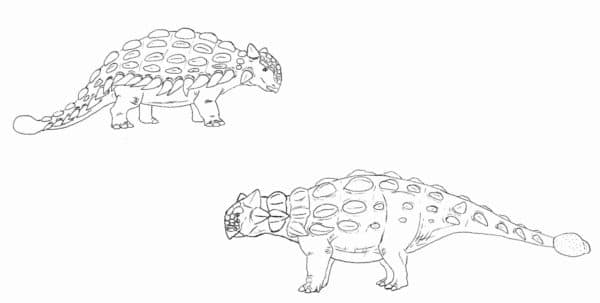
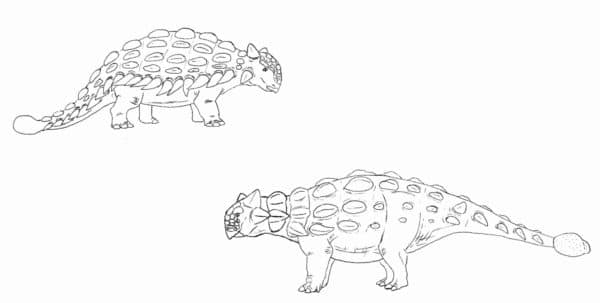
Vintage Ankylosaurus compared to our modern interpretation of the genus. Those lateral spikes weren’t actually there. By Nicholas Carter
Speaking of armor, let’s talk about the Thyreophora, the clade containing the bony scelidosaurs, stegosaurs, and ankylosaurs. Mid-late 20th Century restorations of these animals often tend to emphasize their stout legs, wide bodies, and armor plating. This often resulted in things like drooping, dim-looking stegosaurs with bodies arced in a bell-curve and turtle-like ankylosaurs with sprawling limbs and generic-looking armor. While it’s certainly true that derived stegosaurs and ankylosaurs weren’t the most gracile and light-footed animals, they also weren’t dinosaurian slugs bumbling through the Mesozoic woodlands and surviving on pure luck. Modern discoveries of nicely preserved thyreophorans show that even ‘extreme’ species like Stegosaurus and Ankylosaurus weren’t as stumpy and wilted-looking as was once thought. They had surprisingly longish necks for low-level browsing and the shape and arrangement of their armor and other ornamentation was highly specific to each genus. Some wonderfully fossilized ankylosaurs such as specimens of Edmontonia, Zuul, Sauropelta, and Borealopelta bear armor arranged as it would have been in life, leaving very little to the imagination when it comes to reconstructing these dinosaurs (Brown et al, 2017). Some even had bony plates in the cheeks and eyelids.
Most stegosaurs were built fairly long and low, aside from their protruding plates and spines. The largest, Stegosaurus, could be roughly elephant-sized and had straight columnar limbs to support its weight. Ankylosaurs were very round and expanded in cross-section thanks to their barrel-shaped ribcage. While there’s no direct evidence of this, I like to imagine late Cretaceous ankylosaurs like Panoplosaurus and Scolosarus lounging in swampy bayous during the hot periods like large mammals and tortoises do today.
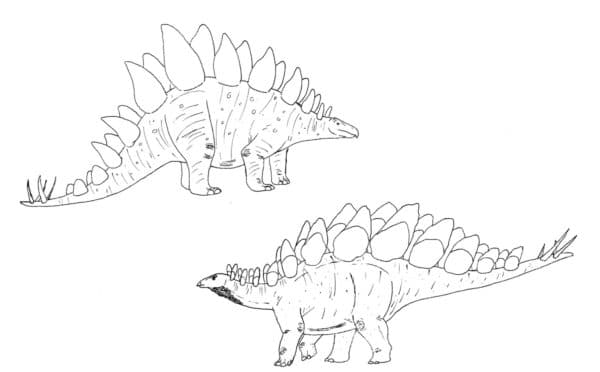
The Stegosaurus of yesterday vs the Stegosaurus of today. By Nicholas Carter
Basal neornithischians came in all sorts of small, fuzzy, scampering forms that dwelled in the undergrowth. Heterodontosaurs, parksosaurs, psittacosaurs… However, I’ll stick to my familiar biases and look most closely at the more derived groups. The iguanodontians were to the Mesozoic world what ungulate grazers and browsers are to today, filling comparable roles to modern sheep, deer, and antelope. Smaller forms were probably bipedal, with larger more derived species switching to a quadrupedal stance, but able to rear up when need be.
The most familiar iguanadontians to us in western North America are the hadrosaurs. These are often nicknamed the ‘duck-billed’ dinosaurs due to their broad, toothless, duck-like beaks. Older depictions of hadrosaurs show the outer soft tissue of the beak matching the shape of the inner bone, and this made sense to early paleontologists who believed hadrosaurs were aquatic and strained water plants with a flat beak. However, exceptionally preserved Edmontosaurus specimens with semi-intact rhampotheca (or ‘horny beak coverings’) show that their beaks were narrow and shovel-like in life (Morris, 1970). We now know that hadrosaurs spent most of their time on land, and this beak design was good for indiscriminately shoveling in whatever vegetation they came across.
We’re blessed with a generous helping of hadrosaur skin impressions from both Asia and North America, which also reveal a good amount about these creatures appearance (Bell, 2014). We know from ‘mummified’ specimens that regions like the neck and tail were much deeper than just the skeleton would suggest. Hadrosaurs follow the general dinosaurian trend of small, beaded, non-overlapping scales, some of which diversified into larger pointed shapes on the belly or limbs of some taxa. Some regions like the neck and shoulders had ridges, bumps, or vertical folds in the skin that added to the texture of these animals. Frills, ridges, serrations, and spike-like ‘feature scales’ ran along the back of various hadrosaurs as well. We now know of soft-tissue crests in Edmontosaurus regalis at least, and it’s entirely possible that other species had such features that we just haven’t found yet (Bell et al, 2013).
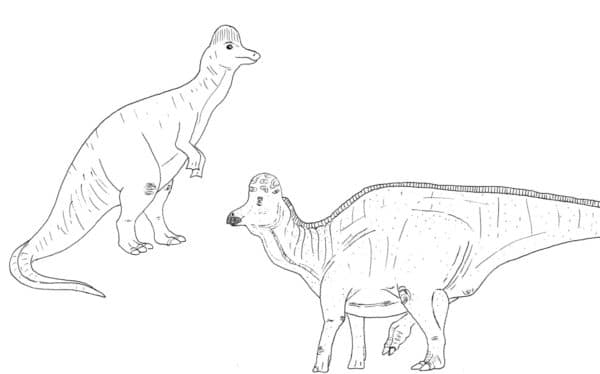
Upright swamp-dweller? Nope, Corythosaurus was a horizontal land animal. By Nicholas Carter
Ceratopsids have also given us a variety of fossilized skin impressions. Some species tended to have fairly regular scale patterns, but some chasmosaurines had large, prominent tubercles surrounded by standard small scales (Sternberg, 1925). This was taken to the extreme in giants like Triceratops, which had protuberant, pointed tubercles that likely would have been noticeable some distance away. One of the defining features of ceratopsids is obviously their famous horns. All we have of these, however, are the bony horn cores, which would have been covered in a sheath of keratin when the animals were alive. It’s long been assumed that the horn sheathes simply conformed to the shape of the bony core, while adding some extra width and length to the horn. However, since we know that ceratopsid horns changed shape quite a bit as the animals grew, this could mean that the horn sheathe didn’t completely match the bony core at maturity, but to some degree reflected the shape of the horn in over time (Horner & Goodwin, 2006)( Frederickson & Tumarkin-Deratzian, 2014).
Like hadrosaurs and thyreophorans, the ornaments of ceratopsids, namely the shape and size of the frills and their adorning spikes, were specific to each species. An often-overlooked part of the wacky ceratopsid head is the nasal area, which was a huge depression that may have supported some kind of fleshy display structure. These dinosaurs are also weird in that they retained all five fingers on each hand, although only the inner three had hoofs. Aside from the basal ceratopsian Psittacosaurus, we don’t have much of an idea as to what colour horned dinosaurs were (Vinther et al., 2016). While the bodies were likely unremarkable, it would make sense that their frills were brightly coloured at least some of the time.
Horned dinosaurs were commonly restored with sprawling front limbs in the past, and this with an exaggerated back curve gave these reconstructions a very hunched-over look. Modern paleontologists favour a ‘somewhat upright but still sort of sprawled’ stance for these animals, which is supported by what trackways we have from them (Paul & Christiansen, 2000)(Thompson & Holmes, 2007).
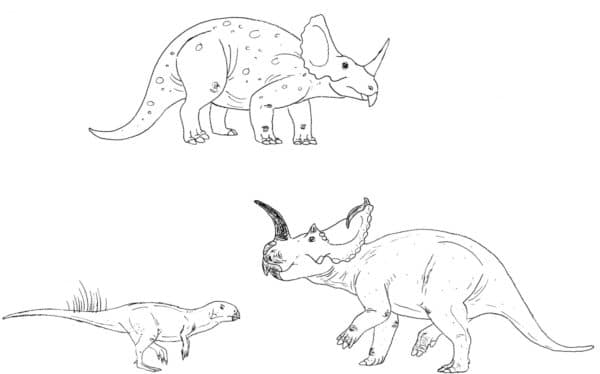
Old-timey Centrosaurs compared to a modern Centrosaurus. Less like an elephant-lizard, more like a rhino-bird. Also the basal ceratopsian Psittacosaurus as we know it today. By Nicholas Carter
Relatively less is known about the life appearance of the dome-headed pachycephalosaurs. This is because, aside from the skull domes, bones of these animals are very rare. Thanks to a few exceptions, like Alberta’s own Stegoceras, we can tell that they were small to medium-sized bipedal dinosaurs with rigid tails and a more horizontal stance than is often portrayed in the media. One of the many weird things about the pachycephalosaurs is their exceedingly broad ribcages and long transverse processes at the base of the tail, giving their bodies a wide bulge on the sides that promptly narrowed at the neck and tail (Paul, 1987)
Moving on to the biggest of all herbivores, the sauropods, it can be hard to imagine anything this big as a living, breathing creature. Since the largest land animals we have today are elephants, sauropods often get reconstructed with wrinkly, elephant-like skin. This is inaccurate, however, since we have skin impressions from these creatures as well. For such massive animals, sauropods had very small scales which would have been extremely hard to make out visually unless you were standing right next to one of these creatures (Czerkas,. 1994). The only exceptions are some titanosaurs like Saltasaurus which bore plate-like osteoderms, and at least some diplodocids which appear to have had a row of spike-like feature scales at least part way along the back (Czerkas, 1992). Their limbs also used to get shown as either much like that of an elephant or tortoise, but there isn’t really a good living analogue for sauropod limbs. Basal sauropodomorphs were a bit more run-of-the-mill as far as dinosaur limbs go, but derived sauropods took things in a weird direction. Like elephants, their legs were held straight in order to support the animal’s bulk. But the feet were a different story altogether. The front feet lacked toes except for a single claw, with the long bones of the hand forming a sort of fleshy, weight-bearing ‘horseshoe’. As for the hind feet, sauropods walked on their toes, with a fleshy pad behind providing cushioning. The hind claws curved out to the sides in a bizarre manner, making sauropod tracks all the more distinctive.
Theories on head and neck position in sauropods has changed over time, with the current idea being that the head and neck, in a neutral pose, were held upright (though, obviously, they could certainly have struck other poses when need be)(Taylor et al, 2009). Many sauropods like apatosaurs and camarasaurs had extremely broad, chunky neck vertebrae, so forget any visions you’ve had of these dinosaurs with curling, swan-like necks. The narial openings of many sauropods are positioned well back on the head, something which has led paleontologists to theorize that they had strikingly posterior nostrils or, even more bizarrely, a trunk. This was used to support the aquatic sauropod idea, with the high-placed nostrils acting like some sort of snorkel. In reality, the nostrils of sauropods were almost certainly placed at the end of the snout, with fleshy passages leading back to the top of the skull. The anatomy and trackways of sauropods all tell us that they were firmly terrestrial animals.
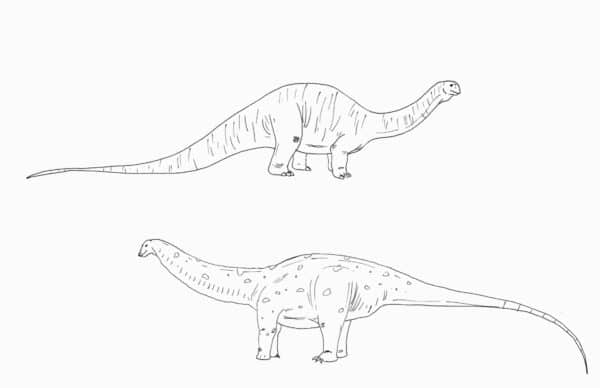
What we know about Brontosaurus has also changed a lot. Lost some bulk off the head, but check out that chunky neck. By Nicholas Carter
Theropods were an extremely diverse group, occupying all sorts of shapes, sizes, and ecological roles. They were also animals, not the snarling, drooling movie monsters as often depicted. Many were certainly large, terrifying predators, though others were smaller, feathered, and birdlike. They walked delicately like large ground birds more than stomping, staggering Godzillas. The hips were normally parallel to the ground, with the head upright on an s-curved neck, and the tail held out for balance. Feathered species bore a variety of feather types, with more basal groups covered in downy, hair-like ‘proto-feathers’ while more derived families acquired full-on vaned plumage. Some theropods were as extensively feathered as modern birds, with the plumage extending as far as the ends of the toes and forming an aeroshell over the body. Many species had forelimb feathers extensive enough to form ‘wings’ of some kind, and some like the microraptorines had ‘wings’ on their hindlegs too. Feathers also formed crests, tail fans, rudders, and other such structures.
Species that had scales generally lacked large scutes or osteoderms except for at least some ceratosaurs, which appeared almost armored. Even large non-feathered tyrannosaurs had fairly small, regular scales. Given the powerful jaw closing capabilities of some species like Tyrannosaurus, many theropods likely had big, muscular jaws and necks to power these killing tools. Despite the iconic toothy grin often given to theropod dinosaurs, the surface texture of their jaw bones indicate that they likely had lips of some kind that would have covered their teeth when the mouth was closed, and thick gums and other oral tissues likely helped conceal much of the teeth. That is, unless it was a beaked theropod. Toothless beaks arose several times in these dinosaurs, and most species that had them seem to have been herbivores. Keratinous rhamphotheca just like that of birds have even been found in theropods like ornithmoimids (Norell et al, 2001). The word theropod means ‘beast-foot’. Ironically, their feet were generally much more like that of birds, with 3 functional toes and perhaps tarsal scutes in some species. Also like birds, they had thick fleshy pads along their toes for weight support. The sadly occupied Ornithopoda would have been a more fitting name for these dinosaurs.
Theropods are really hard to sum up as a group thanks to their wide variety of shapes and sizes, which makes it all the more puzzling when they get depicted as just slight variations of the generic old ‘move monster’ trope. Different lineages sported crests, nasal horns, orbital hornlets, and bizarre sails. While larger species may have been coloured fairly dull so as to not be seen by their prey, these wacky structures were probably quite colourful at least some of the time.
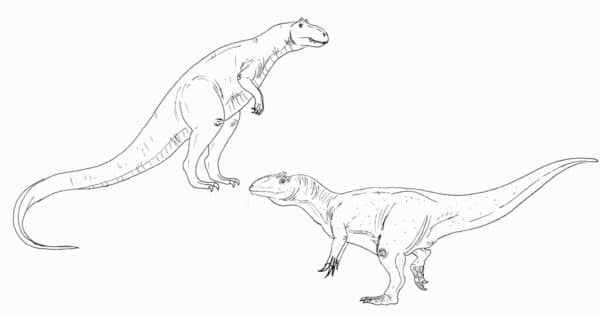
Allosaurus wasn’t some hunched-over kangaroo-lizard. It was agile, sleek, and powerful. By Nicholas Carter
In the interest of not dragging this monster of an article out any further, we’ll draw the line on dinosaur life appearance here for now. Obviously there’s much more that can be said on what these animals, and other prehistoric creatures, looked like when alive. Despite all these, nearly all our best attempts at reconstructing dinosaurs are almost certainly wrong on some level. There’s still a lot of uncertainty in the fine details regarding the appearance of prehistoric animals, and paleontologists can only hope to find more data on this subject. It might be a frustrating notion that we’ll never get a perfect image of live nonavian dinosaurs, but science is always working its way bit by bit towards the truth. We’ve evolved a lot since the early days of paleontology, and in this new golden age of the science, it’s anybody’s guess what we’ll discover next.
By Nicholas Carter
References
Bell, P. R.; Fanti, F.; Currie, P. J.; Arbour, V.M.(2013). “A Mummified Duck-Billed Dinosaur with a Soft-Tissue Cock’s Comb”. Current Biology. 24 (1): 70–75
Bell, P. R. 2014. A review of hadrosaurid skin impressions. In Eberth, D. A. & Evans, D. C. 2014. Hadrosaurs. Indiana University Press, Bloomington & Indianapolis, pp. 572-590.
Brown, C.M.; Henderson, D.M.; Vinther, J.; Fletcher, I.; Sistiaga, A.; Herrera, J.; Summons, R.E. (2017). “An Exceptionally Preserved Three-Dimensional Armored Dinosaur Reveals Insights into Coloration and Cretaceous Predator-Prey Dynamics”. Current Biology. 27 (16): 2514–2521.e3.
Czerkas, S. A. 1992. Discovery of dermal spines reveals a new look for sauropod dinosaurs. Geology 20, 1068-1070.
Czerkas, S. A. 1994. The history and interpretation of sauropod skin impressions. Gaia 10, 173-182.
Frederickson JA, Tumarkin-Deratzian AR. 2014. Craniofacial ontogeny in Centrosaurus apertus. PeerJ 2:e252 https://doi.org/10.7717/peerj.252
Horner, J.R.; Goodwin, M.B. (2006). “Major cranial changes during Triceratops ontogeny”. Proceedings of the Royal Society B: Biological Sciences. 273 (1602): 2757–2761. doi:10.1098/rspb.2006.3643. PMC 1635501. PMID 17015322
Li, Q.; Gao, K.-Q.; Vinther, J.; Shawkey, M. D.; Clarke, J. A.; d’Alba, L.; Meng, Q.; Briggs, D. E. G. & Prum, R. O. (2010). “Plumage color patterns of an extinct dinosaur” (PDF). Science. 327 (5971): 1369–1372.
Morris, William J. (1970). “Hadrosaurian dinosaur bills — morphology and function“. Contributions in Science (Los Angeles County Museum of Natural History) 193: 1–14.
Nabavizadeh, A. (2019), New Reconstruction of Cranial Musculature in Ornithischian Dinosaurs: Implications for Feeding Mechanisms and Buccal Anatomy. Anat Rec. doi:10.1002/ar.23988
Norell, Mark & Makovicky, Peter & Currie, Philip. (2001). Paleontology: The beaks of ostrich dinosaurs. Nature. 412. 873-4. 10.1038/35091139.
Osborn, Henry Fairfield (1909). “The epidermis of an iguanodont dinosaur”. Science. 29 (750): 793–795.
Osborn, Henry Fairfield (1912). “Integument of the iguanodont dinosaur Trachodon”. Memoirs of the American Museum of Natural History. 1: 33–54.
Paul, G. S. 1987. The science and art of restoring the life appearance of dinosaurs and their relatives – a rigorous how-to guide. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present Vol. II. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and London), pp. 4-49.
S Paul, Gregory & Christiansen, Per. (2000). Forelimb posture in neoceratopsian dinosaurs: Implications for gait and locomotion. Paleobiology. 26. 10.1666/0094-8373(2000)026<0450:FPINDI>2.0.CO;2.
Persons, W. S. and Currie, P. J. (2011), The Tail of Tyrannosaurus: Reassessing the Size and Locomotive Importance of the M. caudofemoralis in Non‐Avian Theropods. Anat Rec, 294: 119-131. doi:10.1002/ar.21290
Sternberg, C.M., 1925, “Integument of Chasmosaurus belli”, Canadian Field-Naturalist 39: 108-110
Taylor, M.P., Wedel, M.J. and Naish, D. 2009. Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica 54(2): 213-220.
Thompson, Stefan and Holmes, Robert, 2007. Forelimb Stance and Step Cycle in Chasmosaurus Irvinenesis (Dinosauria: Neoceratopsia). Palaeontologia Electronica Vol. 10, Issue 1; 5A:17p, 8MB; http://palaeo-electronica.org/paleo/2007_1/step/index.htm
Vinther, Jakob; Nicholls, Robert; Lautenschlager, Stephen; Pittman, Michael; Kaye, Thomas G.; Rayfield, Emily; Mayr, Gerard; Cuthill, Innes C. (2016). “3D Camouflage in an Ornithischian Dinosaur” (PDF). Current Biology. 26 (18): 2456–2462.
