
It seems like, back in the good old pre-Dinosaur Renaissance days, the number of dinosaur species known to science was a pretty self-contained thing. You had your Tyrannosaurus, your Triceratops, your Stegosaurus, Brontosaurus, ‘Monoclonius’, and all the other classics. It was kind of like a baseball team of fossil animals. This was never really true, of course, because despite the fact that only a dozen or so animals ever seemed to make it into the popular science books of yesterday, there was hundreds of more obscure dinosaurs that never got much time in the limelight being named at the same time as the star species.
Nowadays, with paleontology being more popular than ever and with people getting better and better at it all the time, new dinosaur species are being named on a weekly basis from all over the world. According to Wikipedia, for non-avian dinosaurs we’re currently sitting at around 500 genera, the first and more inclusive part of an animal’s scientific name, such as Canis for dogs. Within those genera we’ve got around 1000 species. That’s the second and more exclusive part of a scientific name, such as the ‘lupus’ in Canis lupus (the wolf), or ‘latrans’ in Canis latrans (the coyote). But even this number is up for debate among scientists, and the number of dinosaur species still waiting underground for us to find is hard if not impossible to say. This makes the question of how many types of dinosaurs were there a pretty tricky one to answer.
There’s a few different ways of discovering a new genus or species of dinosaur. One way is to go out and dig up something unique that’s never been found before. You might think that nowadays this happens only in relatively unexplored places in the world where paleontologists are just beginning to study the local rocks, and it’s certainly true that underexplored areas are yielding new types of dinosaur. Keeping in Alberta’s borders, the late Cretaceous Wapiti Formation of the northwest, where the Philip J. Currie Dinosaur Museum is located, could be an example of this. There are, however, even in famously well-known areas like the southern half of the province, brand new dinosaurs still pop up from time to time. Sometimes what we find is a new species of an already existing genus, though sometimes it’s a whole new genus unto itself, giving us an exciting new dinosaur name to talk about. Naming a new genus or species from something recently excavated is a time-consuming process, as once the animal is collected it needs to be prepared, carefully studied, subject to a rigorous scientific description, and once the description is accepted and published does everyone have a new dinosaur to enjoy. Despite advances in techniques and technology, paleontology still often moves at a glacial pace.
We also have to remember the fact that only certain slices of time are geologically preserved in certain areas on Earth. We don’t have extensive rock exposures of every age, period, and era from every place in the world to look for fossils in. That means that there are entire chunks of geologic time that we may never be able to explore and know what fossil animals lived at that interval. Whole stages of dinosaur evolution and diversity may forever be a mystery. And to top that off, we can’t expect that members of every species of dinosaur that ever lived were lucky enough to be fossilized. Becoming a fossil requires an animal dying in the right time and place for the environmental conditions to mineralize the bones and other hard parts. The hard truth is that many dinosaurs probably lived in environments that weren’t conducive to the fossilization process, and so the record of their entire existence is gone forever. Isn’t that horribly frustrating? All we can do is continue to dig where the rocks are, and keep on hoping.
Another way of finding a new type of dinosaur is to search in museum or university collections for things that have already been dug up. Paleontology museums all over the world have shelves and drawers and cases full of specimens waiting to be analyzed and described. Paleontologists are ambitious people to be sure, but they can only study so many animals at once. Often times, stuff that’s been sitting around awaiting a good thorough description gets foisted onto graduate students for thesis projects. It’s bound to happen that some unidentified specimens, upon investigation, turn out to be examples of already-established species. Sometimes a partial skeleton gets identified as one type of dinosaur, and then later re-identified as something else after a closer look. An example of this was recently published when what was thought for decades to be a young Daspletosaurus at the Canadian Museum of Nature collected from a very unexpected rock formation was most likely an Albertosaurus from right when we’d expect to find one (Mallon et al., 2019). And sometimes, fossils that are presumed to be from an established genus or species turn out to be something totally new. One example of this (though there are many others out there) is the horned dinosaur Coronosaurus brinkmani named in 2012, which was originally described seven years earlier as a species of Centrosaurus (Ryan & Russel, 2005) (Ryan et al., 2012).
All of this tells us that there’s an indefinite number of dinosaur genera and species in the ground or hiding in museums waiting to be described and named. We aren’t going to run out of new dinosaurs any time soon. However, there’s another reason to be uncertain about the number of dinosaur types out there, and that’s the validity of existing genera and species that, according to some, might not be so secure. When it comes to taxonomy, the science of naming and grouping organisms, there’s two opposing schools of thought. One group are the ‘lumpers’, scientists who place more emphasis on the similarities between slightly different animals, and are more likely to group them together under the same genus or species. The other type are ‘splitters’- those who see more differences between animals and split them into different genera or species. ‘Lumpers’ and ‘splitters’ exist in the taxonomy of both modern animals and extinct ones, and this causes us to think twice about how we name and categorize species.
Species are determined to be distinct from or the same as one another based on a few criteria. Do they look noticeably different, sound different, or behave differently? Is their internal anatomy unique? Perhaps the most important question to ask is, when comparing two similar but possibly distinct animals, is whether they can produce completely fertile young together. If they can, they’re probably the same species. This is the Biological Species Concept, and it’s used as a general rule of thumb by zoologists to determine if animals are the same or different species (it does, however, tend to break down in botany and microbiology). Interbreeding between different species is certainly possible, but tend to either result in infertile hybrids (such as when horses and donkeys cross to produce mules), or offspring that don’t survive beyond shortly after birth, such as in attempted crosses between captive African and Asian elephants.

Pachyrhinosaurus (top right) and Styracosaurus (bottom left) on display at the Philip J. Currie Dinosaur Museum. Their skulls are pretty different, but the rest of their bones are fairly similar. What should we make of this? Photos by Nicholas Carter
At the genus level, things are also complicated. For a long time, things like appearance, behavior, and internal anatomy were used to differentiate species, but with the relatively recent power of genetic sequencing, we can resolve things in better detail. For example, about half of North American ducks were placed in the genus Anas for the past several decades. However, recent molecular analyses have revealed that this is incorrect (Gonzales et al, 2009), and several species were split into genera like Spatula for teals and shovelers and Mareca for wigeons and the Gadwall (Gill & Donsker, 2017). This isn’t to say that there’s no debate about the importance placed on molecular evidence over anatomical data, but we at least have multiple tools to use for figuring out how to classify modern animals.
When it comes to fossil animals, however, there’s much less to go on. Without the benefit of molecular data along with observations on interbreeding, behavior and appearance to go on, the only real tools we have for classifying fossil species are anatomy and relative dating. Paleontologists closely analyze the bones of extinct animals like dinosaurs and gather data on their anatomical traits, or characters, which tells them if they’re looking at different genera/species or not. The rock formation in which fossils are found is also informative, since it’s pretty unlikely that the same species will be found in different formations representing different time spans, but different species in the same genus certainly could be found in slightly different formations. For example, the duck-billed dinosaur Hypacrosaurus sternbergi is found in the Two Medicine Formation, while Hypacrosaurus altispinus is found in the younger Horseshoe Canyon Formation (Horner et al., 2004).
For nearly the entire history of paleontology, there’s been an ongoing debate over the validity of many dinosaur genera and species. Paleontological lumpers see more similarities between different animals, and argue for their grouping together under fewer names, while splitters argue do the opposite. There’s a lot that influences decisions for lumping or splitting dinosaurs. Some fossils might be incomplete, and appear to be unique when they might not be. The multitude of dubious dinosaur genera named after isolated teeth demonstrates this headache nicely. Alternatively, some incomplete specimens appear to belong to already established taxa at first glance, but aren’t complete enough to say for sure. The mystery Dinosaur Park pachyrhinosaur of the Royal Tyrrell Museum is one such example (Ryan et al., 2010).
Discoveries of juvenile dinosaurs over the years also muddy the taxonomic waters. Dinosaur skulls changed quite a bit as the animals grew, especially pertaining to their head ornamentations like crests, horns, and frills. We’re still only just learning how radically dinosaur skulls changed throughout growth. Skulls, or fragments of skulls, from what appeared to be unique species have been re-classified as juveniles or sub-adults of other dinosaurs. Names like ‘Tetragonosaurus’ (Evans et al., 2005), ‘Monoclonius’ (Ryan, 2006), and ‘Brachyceratops’ (Ryan, 2007) have been lumped into other genera typified by adult specimens.

Some of the variation in frills seen in Pachyrhinosaurus lakustai. Dinosaurs certainly weren’t all identical! By Nicholas Carter
We’re also just beginning to figure out how much variation in body shape and size was possible between individuals of the same species in dinosaurs. Paleontologists and paleoartists have long been strict about reconstructing dinosaurs along fairly narrow lines of size and shape. However, we’re figuring out that, just as modern animals of the same species can come in a variety of forms due to genetics, injuries, mutations, and other such factors, so too could dinosaurs. The ceratopsid Pachyrhinosaurus lakustai, pride of the Philip J. Currie Dinosaur Museum, is often reconstructed in art with a fairly symmetrical frill and spike arrangement- the inner two frill spikes or epiossifications being small and pointing inwards, the second pair being long and curving out to the side, and the rest being small nubs. However, the wild variety of fill morphologies dug out of the Pipestone Creek bonebed shows us that this dinosaur’s ornamentation came in all sorts of asymmetrical shapes (Currie et al., 2008). In more recent news, a new complete skull of the dinosaur Styracosaurus has proven that this genus showed a fair degree of variation and asymmetry in its skull too, and the authors took this as a good reason to lump the similar yet enigmatic Rubeosaurus in with Styracosaurus (Holmes et al., 2019).
Sometimes, the reason for dinosaur taxa to be lumped or (more often) split simply comes down to the ego and competitiveness of certain paleontologists. This was demonstrated most clearly during the late 19th century Bone Wars, when rival paleontologists O.C. Marsh and E.D. Cope famously tried spite one another by naming the most species while invalidating the other man’s species. Marsh, for example, named over half a dozen species of Triceratops. Only two are considered valid today. Cope named a handful of genera himself, all quite fragmentary, that most likely belong to Triceratops.
If you look at dinosaur genera, on the other hand, you might notice that most contain only one or two species. Contrast this with genera of living animals, and it’s pretty weird. The lizard genus Varanus alone contains over 50 living species with a wide diversity of sizes and shapes. In the modern extant archosaurs, there’s 14 species in the genus Crocodylus, and genera of modern birds contain anywhere from one to several dozen species. Entomology blows this out of the water, with some insect genera containing over 100 species each. This has led some authors to suggest that dinosaurs are over-split, and that many similar genera should really be lumped together. This has especially pertained to genera that tend to have different-looking skulls but are pretty darn similar from the neck down. This tendency for paleontologists to split genera of fossil animals often comes from a cautious mindset- if you can name something, often best not to assume it’s the same genus as something else. With their variation in size and shape, it seems like dinosaur species put a lot of value in standing out from one another. Other fields of biology tend to focus on the overall similarities, though. Entomologists would likely balk at the idea of the ceratopsids, for example, being over 40 distinct genera with only one or two species in most of them.
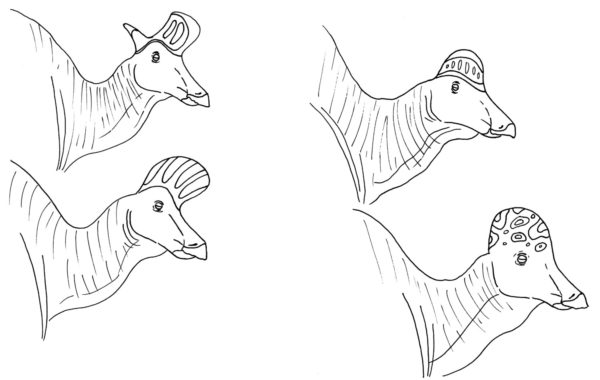
Clockwise from the top left: Lambeosaurus lambei, Hypacrosaurus altispinus, Corythosaurus casuarius, and Lambeosaurus magnicristatus. Are they actually all the same genus? Does it matter? By Nicholas Carter
Author and artist Gregory S. Paul has historically gone to controversial lengths with dinosaur lumping. Arguing that many related dinosaur genera were indistinguishable beyond their skulls, Paul has attempted to synonymize many dinosaur genera over the years. Some well-known cases include following Dale Russell’s suggestion that the tyrannosaurs Albertosaurus and Gorgosaurus are the same genus (Russell, 1970), something that most modern tyrannosaur experts don’t agree with, and the now-abandoned synonymy of Velociraptor and Deinonychus (Paul, 1988). Paul has also argued, for example, that the lambeosaurines Corythosaurus, Lambeosaurus, and Velafrons should be lumped in with Hypacrosaurus, and that a good chunk of centrosaurines should be lumped under Centrosaurus (Paul, 2016). Dr. Darren Naish says more about this here. If these examples (and the many others that Paul has as asserted) were widely accepted by paleontologists, the number of valid dinosaur genera would be remarkably low. However, such a wide taxonomic reworking would require an enormous amount of research and data to be scientifically supported. Subjective observations on the similarity of different animals below the neck don’t make for hard evidence. It’s interesting to note, however, that Paul has also proposed some taxonomic splits in dinosaur genera such as Giraffatitan from Brachiosaurus (Paul, 1988) and Mantellisaurus from Iguanodon (Paul, 2008), which have since seen support from scientists (Taylor, 2009) (Norman, 2010).
If, by now, you’re starting to wonder what even constitutes a dinosaur genus anymore, well, you’re not alone. In addition, if you’re wondering if it even really matters that much anyway, you aren’t alone either. Naming and categorizing dinosaurs is a useful way of scientifically studying and understanding them. As humans, we love to sort, classify, and count things. Does it really matter, though, if for example Eotriceratops xerinsularis turned out to be just an early species of the genus Triceratops? Does anything really change? Not really. Paleontologists and other dinosaur nerds (like me) love to debate the minutiae of things like names and classifications, but these are all human creations to describe living things at one particular point along an evolutionary spectrum.
So, the number of dinosaur types is not a definite, ascending number. Rather, it partly depends on who you talk to and what evidence you value. Regardless of all the debate though, paleontologists continue to discover new ones, and that will always be exciting.
By Nicholas Carter

References
Currie, P.J., Langston, W., and Tanke, D.H. (2008). “A new species of Pachyrhinosaurus (Dinosauria, Ceratopsidae) from the Upper Cretaceous of Alberta, Canada.” pp. 1-108. In:
Currie, P.J., Langston, W., and Tanke, D.H. 2008. A New Horned Dinosaur from an Upper Cretaceous Bone Bed in Alberta[dead link]. NRC Research Press, Ottawa, Ontario, Canada. 144 pp.
Evans, David C.; Forster, Catherine F.; Reisz, Robert R. (2005). “The type specimen of Tetragonosaurus erectofrons (Ornithischia: Hadrosauridae) and the identification of juvenile lambeosaurines”. In Currie, Phillip J.; Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press. pp. 349–366.
Gill, Frank; Donsker, David, eds. (2017). “Screamers, ducks, geese & swans”. World Bird List Version 7.3. International Ornithologists’ Union. Retrieved 23 July 2017.
Gonzalez, J.; Düttmann, H.; Wink, M. (2009). “Phylogenetic relationships based on two mitochondrial genes and hybridization patterns in Anatidae”. Journal of Zoology. 279: 310–318.
Robert.B. Holmes; Walter Scott Persons; Baltej Singh Rupal; Ahmed Jawad Qureshi; Philip J. Currie (2019). “Morphological variation and asymmetrical development in the skull of Styracosaurus albertensis”. Cretaceous Research. in press: Article 104308. doi:10.1016/j.cretres.2019.104308.
Horner, John R.; Weishampel, David B.; Forster, Catherine A (2004). “Hadrosauridae”. In Weishampel, David B.; Dodson, Peter; Osmólska Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 438–463.
Mallon, J.C., Bura, J.R., Schumann, D. and Currie, P.J. (2019), A Problematic Tyrannosaurid (Dinosauria: Theropoda) Skeleton and Its Implications for Tyrannosaurid Diversity in the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta. Anat Rec. doi:10.1002/ar.24199
Norman, David. (2010). “A taxonomy of iguanodontian (Dinosauria: Ornithopoda) from the lower Wealden Group (Cretaceous: Valanginian) of southern England” (PDF). Zootaxa. 2489: 47–66.
Paul, Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. p. 464. ISBN 978-0-671-61946-6.
Paul, G.S. (1988). “The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world’s largest dinosaurs”. Hunteria. 2 (3): 1–14.
Paul, Gregory S. (2008). “A revised t
axonomy of the iguanodont dinosaur genera and species”. Cretaceous Research. 29 (2): 192–216.
Paul, Gregory S. (2016). The Princeton Field Guide to Dinosaurs 2nd Edition. United States of America: Princeton University Press.
Russell, Dale A. (1970). “Tyrannosaurs from the Late Cretaceous of western Canada”. National Museum of Natural Sciences Publications in Paleontology. 1: 1–34.
Ryan, M.J. (2006). “The status of the problematic taxon Monoclonius (Ornithischia: Ceratopsidae) and the recognition of adult-sized dinosaur taxa”. Geological Society of America Abstracts with Programs. 38 (4): 62.
Ryan, M. J.; Russell, A. P. (2005). “A new centrosaurine ceratopsid from the Oldman Formation of Alberta and its implications for centrosaurine taxonomy and systematics”. Canadian Journal of Earth Sciences. 42 (7): 1369. doi:10.1139/e05-029
Ryan, Michael J.; Holmes, Robert; Russell, A.P. (2007). “A revision of the late Campanian centrosaurine ceratopsid genus Styracosaurus from the Western Interior of North America” (PDF). Journal of Vertebrate Paleontology. 27 (4): 944–962.
Ryan, M. J.; Eberth, D. A.; Brinkman, D. B.; Currie, P. J.; Tanke, D. H., “A New Pachyrhinosaurus-Like Ceratopsid from the Upper Dinosaur Park Formation (Late Campanian) of Southern Alberta, Canada”, in Ryan, M. J.; Chinnery-Allgeier, B. J.; Eberth, D. A., eds. (2010), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium, Indiana University Press
Ryan, M. J.; Evans, D. C.; Shepherd, K. M.; Sues, H. (2012). “A new ceratopsid from the Foremost Formation (middle Campanian) of Alberta”. Canadian Journal of Earth Sciences. 49 (10): 1251. doi:10.1139/e2012-056
Taylor, M.P. (2009). “A Re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropod) and its generic separation from Giraffatitan brancai (Janensch 1914)” (PDF). Journal of Vertebrate Paleontology. 29 (3): 787–806.