To many people, mammals seem like very modern animals. Palaeontologists of the mid-20th Century often framed the story of life on Earth into overly-simplified chapters, with the earlier Paleozoic and Mesozoic eras being the ‘Age of Fish’ and ‘Age of Reptiles’ respectively, and the current Cenozoic being the ‘Age of Mammals’. This might make for easy mental categorization, but it’s also misleading. Mammals themselves date back to the time of the dinosaurs, and their earlier cousins, the non-mammalian synapsids, go back earlier.
While the evolution of mammals isn’t as glamorous in the eyes of the public as dinosaurs, palaeontologists have always been highly interested in fossil mammals and their relatives. When the study of dinosaurs became less popular during the first half of the 20th Century, mammal palaeontology was treated with much weight and seriousness. Today, it’s still a very active field of study, with work being done all over the world to fill in the history of mammals. Not only does this study inform us about where the furry things we share the planet with today came from, it also traces our own ancestry back to the earlier days of the Earth.
Mammals belong to a larger group of animals called synapsids. These are vertebrate animals that produce amniotic eggs and have a single hole, or temportal fenestra, behind their eye socket. The sister group to the synapsids are the sauropsids, all modern ‘reptiles’ (including birds) and their fossil relatives. Many fossil animals might look somewhat ‘reptilian’, but actually belong to the synapsid branch. These animals are sometimes casually called ‘mammal-like reptiles’, but since the term ‘reptile’ is fairly convoluted now, the ‘mammal-like reptile’ term is no longer used by palaeontologists. Instead we use terms like ‘stem-mammals’ or ‘non-mammalian synapsids’.
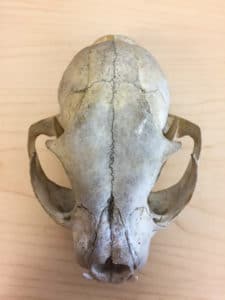
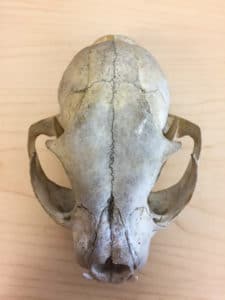
A Canada Lynx skull seen from above. You can see the temporal fenestrae on each side through the semi-open eye sockets above. Photo by Nicholas Carter
Synapsids and sauropsids split off from each other about 312 million years ago, during the late Carboniferous period. One of the earliest known synapsids was the small insect-eating Archaeothyris of Nova Scotia (Reisz, 1972). Archaeothyris looked vaguely like a lizard, but many anatomical features such as the single skull opening behind the eye prove it to be an early synapsid.
Our synapsid forbearers first came to real prominence during the Permian period, a time that began almost 300 million years ago, long before the first dinosaurs. Before that time, in the Carboniferous, the world had been dominated by rainforests, and the distant ancestors of amphibians were the most common land animals. As the separate landmasses converged into the supercontinent Pangea, Earth saw a fair degree of drying, especially away from the coastlines. Inland regions, separated from precipitation from the ocean, turned into arid plains and deserts. In the wake of this rainforest collapse, amphibian-like tetrapods, tied to the water for survival, became more localized, while animals that could lay water-containing shelled eggs (the amniotes, as we call them) saw a boost in diversity (Sahney et al., 2010). This includes the aforementioned sauropsids, who would branch into some successful groups in the Permian but really come into their own in the later Mesozoic, and our synapsid ancestors.
In the early Permian period, synapsids radiated in diversity, taking on some pretty strange forms that dazzled early palaeontologists. Some, like the varanopids and eothyridids, continued to look like somewhat lizard-like, not growing terribly large and likely having a carnivorous/insectivorous diet. A varanopid called Ascendonanus was even recently described with lizard-like scaly skin preserved around the fossil (Spindler et al., 2018). This would indicate that scales are the likely ancestral covering to synapsids as opposed to naked skin or hair. However, if varanopids are actually diapsids and not synapsids as has been newly proposed (Ford & Benson, 2020), the remarkably preserved Ascendonanus may not tell us much about the outer appearance of early synapsids.
There was also the caseids, a family that may have been aquatic and fed on plants. Some species in this family, such as the hippo-sized Cotylorhynchus of the United States, were truly bizarre, with a tiny plant-eating head on a large, barrel-shaped body. A recent study suggests that caseids may have had a simple diaphragm muscle similar to that of modern mammals to assist in their breathing (Lambertz et al., 2016).

The sail-backed Permian herbivore Edaphosaurus. By Nicholas Carter
Other decent-sized herbivores during this time were the edaphosaurids, one of the more familiar species being the family’s namesake Edaphosaurus of the American southwest. Like the caseids, they had small heads on short necks and large, round bodies. They differed, though, in likely being terrestrial and often bearing highly elongated neural spines along the back that likely formed a sail. Unlike in many other sail-backed animals, though, Edaphosaurus had bony ‘crossbeams’ sticking out sideways from each neural spine. The function of sails in animals like this, and the bony crossbeams of Edaphosaurus, is still being debated by palaeontologists. Edaphosaurus was described by early American palaeontologist Edward Drinker Cope (Cope, 1882), who confused the post-cranial bones of this animal for a different species that he believed was similar to his other sail-backed animal, Dimetrodon (more on that one later). Amusingly, Cope believed that the unusual spines of what we now know to be Edaphosaurus formed a series of forward-facing sails along the animal’s back, which he thought it used to cruise around Permian lakes with help from the breeze (Cope, 1886). Whatever the sail of Edaphosaurus was really for, it almost certainly wasn’t this.
Where there are big herbivores, there will be predators to feed on them. Some of the earlier carnivorous synapsids were the sphenacodontids. Some, like Sphenacodon, had short, deep skulls bearing large piercing teeth and a distinct ‘ridge’ or short sail along the back. Later species, like the famous Dimetrodon, were the top predators of their ecosystem and grew tall, elaborate sails similar to that of edaphosaurids. Dimetrodon has become something of a figurehead for non-mammalian synapsids, and over the years has been one of the few species of this group often featured in pop culture palaeontology media. This had led to the sadly still common misconception that Dimetrodon is a type of dinosaur, when really it falls nowhere near dinosaurs in the vertebrate tree of life and is more closely related to you and I. Dimetrodon inhabited what was, at the time, likely a subtropical swamp where it was one of the more dominant predators (Olson, 1966).
There are over a dozen named species of Dimetrodon so far, making it one of the most diverse genra of early synapsids. Most of these species come from early Permian beds in Texas and Oklahoma, and range in size and shape from the lizard-sized Dimetrodon teutonis to the much larger Dimetrondon angelensis. Canada actually has its own species of Dimetrodon too- Dimetrodon borealis from Prince Edward Island. This species, represented by a piece of jaw bone with teeth, was first described by Joseph Leidy in 1854, under the name Bathygnathus borealis. Leidy actually thought this jaw came from a dinosaur, believing to be the first instance of these animals in Canada (Spalding, 1995). After the discovery of sphenacodont synapsids by Cope decades later, it was eventually realized that Bathygnathus was really an animal from that group, but no one really paid much attention to it. Until, in 2015, Canadian palaeontologist Kirstin Brink and colleagues re-described the jaw fragment and proved that it actually came from what had really been a type of Dimetrodon all along (Brink et al., 2015).

Dimetrodon, perhaps the most famous pre-Mesozoic fossil animal. By Nicholas Carter
Just how mammal-like in their metabolism and lifestyle these early synapsid groups were is still something of a mystery. Palaeontologists have long been uncertain as to whether or not these animals were ‘warm-blooded’, and where exactly certain very mammalian traits like hair and specialized skin glands (including mammary glands) arose. As with birds, the traits that make mammals unique in the context of living groups didn’t arise all at once, but bit by bit over time. One thing we’re realizing more and more is that endothermy (or ‘warm-bloodedness’, the ability to regulate one’s body temperature) isn’t a black-and-white thing where you’re either endothermic or not, but rather it’s more like a metabolic spectrum, and some groups contain species that fall on different points along this continuum of energy generation.
It’s starting to seem as though early synapsids may have been not quite cold-blooded, but not quite warm-blooded either. The species Ophiacodon of the late Carboniferous-early Permian has been seen to possess fast-grown fibrolammellar bone, a trait also seen in endothermic birds and mammals. Mammals have cartilaginous structures in their noses nasoturbinals which help to warm and moisten incoming air, and Dimetrodon appear to have had very simple nasoturbinals as well (Kemp, 2006). The degree to which these animals could maintain their own body temperature is still hard to tell and probably varied between different species. Things go especially up in the air when you stop and think about the sails that many of these early synapsids like Edaphosaurus and Dimetrodon had on their backs. Palaeontologists continue to debate whether these were devices for display, as many weird, otherwise unexplainable features of fossil taxa seem to be, (Tomkins et al., 2010) or thermoregulation, acting like big solar panels to warm the blood (Haack, 1986) (Florides et al., 1999). There’s arguments for and against both cases, and we won’t get into all that here. Suffice it to say, the early synapsids were weird, and we’re still figuring out a lot about them.
In future entries in this series, we’ll look at a major clade that split off from a common ancestor with the sphenacodontids, a group called the therapsids, and all the bizarre forms these animals came in. Then, we’ll actually get to the mammals themselves and look at the different groups that have come and gone before today.
By Nicholas Carter

References
Kirstin S. Brink et al. 2015. Re-evaluation of the historic Canadian fossil Bathygnathus borealis from the Early Permian of Prince Edward Island. Canadian Journal of Earth Sciences 52 (12): 1109-1120; doi: 10.1139/cjes-2015-0100
Cope, E. D. (1882). “Third contribution to the History of the Vertebrata of the Permian Formation of Texas”. Proceedings of the American Philosophical Society. 20: 447–474.
Cope, E. D. (1886). “The long-spined Theromorpha of the Permian epoch”. American Naturalist. 20: 544–545.
Florides, G.A.; Wrobel, L.C.; Kalogirou, S.A.; Tassou, S.A. (1999). “A thermal model for reptiles and pelycosaurs”. Journal of Thermal Biology. 24 (1): 1–13
Ford, D.P., Benson, R.B.J. The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae. Nat Ecol Evol 4, 57–65 (2020). https://doi.org/10.1038/s41559-019-1047-3
Haack, S.C. (1986). “A thermal model of the sailback pelycosaur”. Paleobiology. 12 (4): 450–458.
Kemp, T.S. (2006). “The origin and early radiation of the therapsid mammal-like reptiles: a palaeobiological hypothesis”. Journal of Evolutionary Biology. 19 (4): 1231–1247.
Lambertz, M.; Shelton, C.D.; Spindler, F.; Perry, S.F. (2016). “A caseian point for the evolution of a diaphragm homologue among the earliest synapsids”. Annals of the New York Academy of Sciences. 1385 (1): 3–20. doi:10.1111/nyas.13264
Olson, E.C. (1966). “Community evolution and the origin of mammals”. Ecology. 47 (2): 291–302.
Reisz, Robert. (1972). Pelycosaurian reptiles from the Middle Pennsylvanian of North America. Bulletin of the Museum of Comparative Zoology of Harvard. 144.
Sahney, S., Benton, M.J. & Falcon-Lang, H.J. (2010). “Rainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica”. Geology. 38 (12): 1079–1082.
Society of Vertebrate Paleontology. (2015, October 29). Ancestry of mammalian ‘warm-bloodedness’ revealed. ScienceDaily. Retrieved February 2, 2020 from www.sciencedaily.com/releases/2015/10/151029134252.htm
Spalding, D.A.E. (1995). “Bathygnathus, Canada’s first “dinosaur””. In Sarjeant, W.A.S. (ed.) (eds.). Vertebrate Fossils and the Evolution of Scientific Concepts. Taylor & Francis US. pp. 245–254.
Spindler, F., Werneburg, R., Schneider, J.W., Luthardt, L., Annacker, V., & Roessler, R. (2018). First arboreal ‘pelycosaurs’ (Synapsida: Varanopidae) from the early Permian Chemnitz Fossil Lagerstätte, SE Germany, with a review of varanopid phylogeny. PalZ, 92, 315-364.
Tomkins, J.L.; LeBas, N.R.; Witton, M.P.; Martill, D.M.; Humphries, S. (2010). “Positive allometry and the prehistory of sexual selection” (PDF). The American Naturalist. 176 (2): 141–148.