
In late April 2020, a description of the tail vertebrae of Spinosaurus was published in the journal Nature (Ibrahim et al., 2020). Most publications on the slowly-unveiled anatomy of this enigmatic giant theropod tend to get a lot of attention, and shocks. The Ibrahim et al., 2020 paper was par for the course in this, with its assentation that long, delicate spines on the top of Spinosaurus’ tail, features which until recently hadn’t been fully described in this dinosaur, served as an adaptation for swimming.
Despite the unforeseen weirdness of the tail, it wasn’t too surprising to see suggested to be an aquatic propulsion tool. Previous studies have asserted that Spinosaurus was adapted to life in and around fresh water (Ibrahim et al., 2014) (Arden, 2018), though Henderson, 2018 put forth that Spinosaurus wasn’t as hydrodynamic as others have thought, arguing that it spent more time wading in shallow water and along shorelines than outright swimming.
We aren’t here, however, to debate the anatomy and behaviour of Spinosaurus. We’re here to talk about swimming dinosaurs in general. With all its seemingly aquatic adaptations, Spinosaurus often gets thrown around in the popular media and online dinosaur fandom as the lone exception to the rule that dinosaurs were solely land-based animals. Was it really the case that dinosaurs stayed strictly on land, with only Spinosaurus potentially bucking the trend? As with so many other things relating to the behavior of animals, it’s not a clean-cut situation. Fossil animals, just as live ones today, did what they did with no regard for our rules and preconceptions. So is there evidence, or even room for reasonable speculation, that dinosaurs other than Spinosaurus took to the water? Let’s talk about it.
There’s a good reason why, nowadays, dinosaurs are generally shown in art and other media as strictly land-based animals. It’s well-known among those interested in the history of palaeontology that, before the Dinosaur Renaissance of the late 20th century, dinosaurs were normally described by palaeontologists as lazy and sluggish, spending the majority of their time lounging in deep wetlands. This idea came from a few mistaken notions, namely that the teeth and jaws of dinosaurs were too small and weak to process tough land plants, and that dinosaurs were too big to support their own mass out of water for too long, especially sauropods. Throw in the fact that many of the most famous dinosaur fossil localities were deposited in what were then lush, swampy ecosystems, and you have what appears to have been a fairly water-loving group of animals.

The classic and beautiful, yet very inaccurate, depiction of Brontosaurus as lazy, primitive, and swamp-bound from 1897. This was considered an accurate view of dinosaurs over 100 years ago, but could dinosaurs have still entered the water from time to time? Painting by Charles R. Knight, public domain.
This mentality began to change from the 70’s onward, when young palaeontologists argued, correctly, that dinosaurs were generally well-adapted for life on land, and nearly all species showed no evidence for an aquatic or amphibious lifestyle. Studies of herbivorous dinosaur teeth and jaws show that they could and did feed on terrestrial plants, and the posture and limb/foot structure of dinosaurs point to them being best-suited to walking on land.
There are exceptions to this, of course, with the spinosaurs being some of the most famous examples of dinosaurs that spent a fair bit of time in or around water. At least a couple non-avian, non-spinosaurid dinosaurs show skeletal evidence of a semiaquatic lifestyle. The duck-like dromaeosaur Halszkaraptor from Mongolia appears to have swam and fed a bit like modern waterfowl, which is a new and unusual specialization for a dromaeosaur in the fossil record (Cau et al., 2017) (Cau, 2020). What’s even more surprising about this dinosaur is that the ecosystem it inhabited was fairly arid, and has changed relatively little since the late Cretaceous, though was perhaps a little less dry than it is today. So if Halszkaraptor did indeed have a lifestyle similar to that of modern Anseriformes (ducks, geese, swans, etc.), there must have been some fairly consistent freshwater areas for it to occupy.
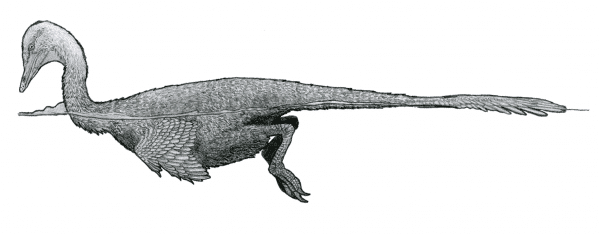
The duck-like dromaeosaur Halszkaraptor bobbing along on the water’s surface. Image: Tom Parker, CC BY 4.0
There’s another possibly aquatic dinosaur that has largely flown under the radar for a while, but is so strange that it deserves more attention in the broader dino-fandom. Liaoningosaurus was a small armored dinosaur, one of the ankylosaurids, from early Cretaceous China. Anyklosaurs were typically large, herbivorous, land-based dinosaurs, but Liaoningosaurus was found with fish bones in its gut, and this combined with its unusual skeletal proportions has led some palaeontologists to think that it may have been aquatic (Ji et al., 2016). Of all dinosaur groups, ankylosaurs arguably seem among the least likely to produce and aquatic specialist species, but here we are…

The potentially semi-aquatic, fish-eating ankylosaur Liaoningosaurus. Image: Jonathan Chen, CC BY 4.0.
Plenty of avian dinosaurs (also known as birds) also specialized in fully or semi-aquatic lifestyles. The poster child for fossil aquatic birds is the nearly 6-foot Hesperornis from Cretaceous North America (Rees et al., 2005). Hesperornis (as well as the many other closely related hesperornithid species) seems to have looked vaguely like a modern loon or grebe, but it’s actually just outside the group containing all modern birds. It had no wings, a long toothed beak, and strong paddling legs used for swimming through the waters of the Western Interior Seaway. Other water-loving birds evolved in the Cretaceous as well, including the early relatives of modern ducks and geese, such as Vegavis from Antarctica (Clarke et al., 2005). And, of course, there’s a myriad of aquatic birds around today, from waders like flamingoes and shorebirds to freshwater divers like loons and grebes to the almost fully marine penguins.
(A side note- one of my palaeontologist colleagues once posed to me the following questions: “Do you consider birds to be ‘reptiles’ in a loose sense?” “Yes,” I responded with bemusement. “And do you consider penguins to be more or less marine animals?” “Yes as well”, though I’m not a penguin ecology expert whatsoever. “Then do you consider penguins to be marine reptiles?” That really opened my mind a fair bit. I supposed I would consider penguins to be marine reptiles in the same manner as plesiosaurs or mosasaurs, even though it just doesn’t feel right for some reason…)
So there certainly were and are dinosaurs that specialized in spending a lot of time in water. What about types that we normally regard as strictly terrestrial, though? Is there any evidence that they took the odd dip in a river, lake, or wetland? Looking at modern animals, it’s far from an unlikely idea. There are a variety of creatures around today that we normally think of as mostly land-based but enter the water on an infrequent or even regular basis.
Large tropical mammals such as elephants, Indian rhinos, and buffalos frequently lounge in shallow pools and rivers, and moose get a good deal of their food outside of winter time from freshwater aquatic plants. Carnivores as well, such as bears, tigers, and jaguars enter the water often and willingly to cool off or catch food. Some monitor lizards will go into the water on a regular or semi-regular basis, and some large tortoises appear to enjoy lounging in cool shallow pools. A lot of this behavior would never be obvious to us if we knew these animals only from skeletal remains alone. We’d have no way of knowing just by looking at their bones that tigers, for example, enjoy a regular swim while lions generally avoid the activity. Even in fairly aquatic animals like hippos, you’d need to study their anatomy well to infer their largely freshwater lifestyle.
This opens up a lot of possibilities for unspecialized swimming behavior in non-avian dinosaurs. With such great body masses, you might think that dinosaurs would’ve sank like rocks, but many giant species had a variety of weight-saving adaptations in their bodies which made walking (and perhaps swimming) around easier. Dr. Donald Henderson of the Royal Tyrrell Museum has long been curious about how different types of dinosaurs performed in water. Using computer simulation models, he found that sauropods, the biggest land animals that ever lived, actually floated pretty nicely (Henderson, 2004). Some sauropods even appeared to be extra-buoyant from the hips backwards, which would’ve left their posterior ends floating and their front limbs pointing downwards. These results may compliment findings of sauropod trackways composed of only the front feet of the animals (Vila et al., 2005), though the authors of this trackway study are skeptical.
Henderson’s models also found that hadrosaurs seemed to have floated in water with their heads just above the surface, meaning they wouldn’t have struggled much to breathe in calm circumstances (Henderson, 2014). It was also found, though, that while ceratopsids would’ve floated too, it was in a head-down position, which would’ve made them poorly-suited to being in water of any significant depth. This might help explain the abundance of vast horned dinosaur bonebeds in Alberta of species like Centrosaurus, Styracosaurus, and Pachyrhinosaurus– when caught in violent flash-floods, these dinosaurs may have been predisposed to drowning easier than other types of dinosaurs, especially in panicked herds of hundreds or even thousands of animals.
Multiple pieces of trackway evidence shows that at least some theropods could swim, and did so on occasion (Coombs, 1980) (Ezquerra et al., 2007) (Xing et al., 2013). With their hollow bones, air sac system, and powerful hind limbs, it seems like all but the largest and bulkiest theropods would’ve done a respectable job of swimming when the need called for it. Why would non-aquatic theropods have needed to swim in the first place, though? There could be many reasons why, but I suspect it was for often mundane reasons, such as crossing a river to get to prey on the other side.
There’s a possibility that some dinosaurs may have even waded in shallow water regularly. In places like southeastern Alberta, swampy areas with lots of shallow water and mud would’ve been common on the landscape, and some dinosaurs may have waded like modern shorebirds, herons, and rails through the wetlands to seek shelter and food. Caenagnathids, for example, might have been specialized for wading through marshy areas based on the structure of their pelvis and long hind limbs (Currie & Russel, 1988). No doubt, many other dinosaurs would’ve had to content with wet, swampy ground from time to time as well.
It probably wouldn’t surprise most palaeontologists if many dinosaurs that lived in warmer, wetter places spent time lounging in shallow pools or rivers in the same manner as modern elephants, rhinos, and water buffalo. In places like Cretaceous Alberta, larger dinosaurs would’ve been at risk of overheating during the summer, and likely needed ways to bring their body temperatures down to a tolerable range after moving around in the heat (O’Connor and Dodson, 1999). Perhaps some, or even most, species habitually lounged about in shallow water or mud wallows during warmer parts of the day.
A bit more conjectural, however, is the idea that some dinosaurs may have lived semiaquatic, hippo-like lives, spending much of day time shielded from the heat of the sun below the water’s surface and emerging in the cool nighttime to feed. Such a lifestyle has been suggested for horned dinosaurs, most notably the relatively long-necked and robust Anchiceratops (Langston, 1959) (Mallon et al., 2012). Henderson’s 2014 paper, however, contradicts this, as does the observation that the eyes and nostrils of hippos are situated high on the head for looking and breathing above the surface, traits that are absent in ceratopsids.

Plenty of modern dinosaurs enjoy hanging out in water, such as this Horned Grebe (Podiceps auritus). Photo by Nicholas Carter
So, as you can hopefully see, a dinosaur needn’t be a fin-bearing, crocodile-headed, web-footed spinosaur in order to spend a notable time in or around water. While not strictly confined to the primordial swamps by their sheer bulk as in so many vintage palaeontology works of art, dinosaurs were every bit as varied as modern animals in their behavior and ecology, meaning that at least some almost certainly enjoyed a cool dip or more. Heck, some alien scientist looking at human anatomy would never say that we have any inclination toward entering water, and many of our closest relatives tend to avoid it when possible, but try telling that to a poolside 10-year-old on a summer day.
By Nicholas Carter

References
Arden, T.M.S.; Klein, C.G.; Zouhri, S.; Longrich, N.R. (2018). “Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in Spinosaurus“. Cretaceous Research. 93: 275–284.
Cau, A.; Beyrand, V.; Voeten, D.; Fernandez, V.; Tafforeau, P.; Stein, K.; Barsbold, R.; Tsogtbaatar, K.; Currie, P.; Godrfroit, P. (6 December 2017). “Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs”. Nature. 552 (7685): 395–399.
Cau A. (2020) The body plan of Halszkaraptor escuilliei (Dinosauria, Theropoda) is not a transitional form along the evolution of dromaeosaurid hypercarnivory. PeerJ 8:e8672
Clarke, J.A.; Tambussi, C.P.; Noriega, J.I.; Erickson, G.M.; Ketcham, R.A. (2005). “Definitive fossil evidence for the extant avian radiation in the Cretaceous” (PDF). Nature. 433 (7023): 305–308.
Coombs, W.P. “Swimming Ability of Carniverous Dinosaurs.” Science. Vol. 207. 3/14/1980.
Currie, P. J. & Russel, D. A. (1988) “Osteology and relationships of Chirostenotes pergracilis (Saurischia, Theropoda) from the Judith River (Oldman) Formation of Alberta, Canada” Canadian Journal of Earth Sciences, 1988, 25:972-986, https://doi.org/10.1139/e88-097
Ezquerra, Doublet, Costeur, Galton, Perez-Lorente. 2007. Were non-avian theropod dinosaurs able to swim? Supportive evidence from an Early Cretaceous trackway, Cameros Basin (La Rioja, Spain). Geology 35: 507-510.
Henderson, D.M. (2004). Tipsy punters: sauropod dinosaur pneumaticity, buoyancy and aquatic habits. Proceedings of the Royal Society of London. Series B: Biological Sciences, 271, S180 – S183.
Henderson, D.H. (2014). Duck soup: The floating fates of hadrosaurs and ceratopsians at Dinosaur provincial park. In Eberth, D. A. & Evans, D. C. 2014. Hadrosaurs. Indiana University Press, Bloomington & Indianapolis, pp. 459 – 465.
Henderson, D.M. (2018). “A buoyancy, balance and stability challenge to the hypothesis of a semi-aquatic Spinosaurus Stromer, 1915 (Dinosauria: Theropoda)”. PeerJ. 6: e5409. doi:10.7717/peerj.5409. PMC 6098948. PMID 30128195.
Ibrahim, Nizar; Sereno, Paul C.; Dal Sasso, Cristiano; Maganuco, Simone; Fabri, Matteo; Martill, David M.; Zouhri, Samir; Myhrvold, Nathan; Lurino, Dawid A. (2014). “Semiaquatic adaptations in a giant predatory dinosaur”. Science. 345 (6204): 1613–6. Bibcode:2014Sci…345.1613I. doi:10.1126/science.1258750
Ibrahim, Nizar; Maganuco, Simone; Dal Sasso, Cristiano; Fabbri, Matteo; Auditore, Marco; Bindellini, Gabriele; Martill, David M.; Zouhri, Samir; Mattarelli, Diego A.; Unwin, David M.; Wiemann, Jasmina (2020). “Tail-propelled aquatic locomotion in a theropod dinosaur”. Nature: 1–4. doi:10.1038/s41586-020-2190-3. ISSN 1476-4687.
Ji Q., Wu X., Cheng Y., Ten F., Wang X., and Ji Y. 2016. Fish-hunting ankylosaurs (Dinosauria, Ornithischia) from the Cretaceous of China. Journal of Geology, 40(2).
Langston, W.J. (1959). “Anchiceratops from the Oldman Formation of Alberta”. National Museum of Canada Natural History Papers. 3: 1–11.
Mallon, Jordan C.; Holmes, Robert; Eberth, David A.; Ryan, Michael J.; Anderson, Jason S. (2012). “Variation in the skull of Anchiceratops (Dinosauria, Ceratopsidae) from the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta”. Journal of Vertebrate Paleontology. 31 (5): 1047–1071. doi:10.1080/02724634.2011.601484
O’Connor, M. P., & Dodson, P. (1999). Biophysical constraints on the thermal ecology of dinosaurs. Paleobiology, 25(3), 341-368.
Rees, Jan & Lindgren, Johan; Lindgren (2005). “Aquatic birds from the Upper Cretaceous (Lower Campanian) of Sweden and the biology and distribution of hesperornithiforms”. Palaeontology. 48 (6): 1321–1329.
Vila, Bernat & Oms, Oriol & Galobart, Àngel. (2005). Manus-only titanosaurid trackway from Fumanya (Maastrichtian, Pyrenees): Further evidence for an underprint origin. Lethaia. 38. 211-218. 10.1080/00241160510013312.
Xing L D, Lockley M G, Zhang J P, et al. A new Early Cretaceous dinosaur track assemblage and the first definite non-avian theropod swim trackway from China. Chin Sci Bull, 2013, 58: 2370-2378, doi: 10.1007/s11434-013-5802-6