
It’s safe to say that Parasaurolophus is easily one of, if not the most, popular hadrosaurs amongst the general public. It’s as iconic as it is weird looking, with that long, backward-curving, tube-shaped crest. It’s an almost alien creature in a way. Despite its popularity though (which seems to have arisen around the time of the dinosaur’s bit-part in Jurassic Park), Parasaurolophus remains a relatively rare hadrosaur compared to other famous duckbills from Cretaceous North America, and many strange ideas have surrounded it over the decades. Let’s take a look at what the latest science has to say on the real Parasaurolophus.
The holotype specimen of this dinosaur was found in Dinosaur Provincial Park in 1920 by a team from the Royal Ontario Museum. Levi Sternberg, part of the legendary Sternberg dinosaur hunting family, oversaw the collection of the dinosaur in the field, as well as its mounting back at the museum. The new duckbill was described and named by the Canadian palaeontologist William Parks soon after, who gave it the full name of Parasaurolophus walkeri (Parks, 1922).
The name Parasaurolophus means ‘near (or similar to) Saurolophus’, another species of hadrosaur from the Horseshoe Canyon Formation of Alberta. As written in his description of the dinosaur, Parks believed that the long crest of Parasaurolophus indicated that it was closely related to Saurolophus, and so gave it a name that reflected this belief*. Despite Parks’ skill as a palaeontologist, we now know he was actually mistaken on this front. Saurolophus did indeed have a long, pointed crest, but it was much shorter and more pointed than that of Parasaurolophus. Parks knew this, but saw a closer similarity to his new dinosaur in Saurolophus than the helmet-shaped crests of species like Corythosaurus or Lambeosaurus. However, the problem that really makes Parasaurolophus a misnomer is that the crest of that species is full of winding, hollow tubes inside, just as in other hollow-crested duckbills like the aforementioned Corythosaurus and Lambeosaurus. These species with hollow spaces in their crests make up the hadrosaur subfamily Lambeosaurinae. The crest of Saurolophus, on the other hand, is a solid structure. This makes it a member of the solid-crested (or, sometimes, non-crested) Saurolophinae along with species like Edmontosaurus and Prosaurolophus.
*Side note- many people pronounce Parasaurolophus as ‘Para-sore-all-uff-us. I maintain this is technically incorrect, as the name is derived from Saurolophus, which is pronounced ‘Sore-oh-loaf-us’, and therefore Parasaurolophus should be ‘Para-sore-oh-loaf-us’.
The Parasaurolophus walkeri holotype at the Royal Ontario Museum. Photo by Nicholas Carter.
The function of the wild headgear in Parasaurolophus and its relatives had palaerontologists scratching their heads for decades. In agreement with Barnum Brown’s suggestion regarding Saurolophus, Parks thought that the crest of Parasaurolophus anchored some sort of fleshy, basilisk-style frill that ran along the top of the animal’s neck and back for some distance. For reasons we’ll discuss later, he backed this up with what he thought to be evidence of a ligament that connected the crest to the dorsal vertebrae.
Other palaeontologists published theories on the function of the crest in Parasaurolophus, and hadrosaurs in general, over the years. Since these animals were long thought to be mostly aquatic, Romer suggested that the tube-shaped crest of Parasaurolophus may have been a snorkel to breathe with while feeding in shallow water (Romer, 1933), and other palaeontologists echoed similar ideas related to breathing during submerged foraging (Sternberg, 1935) (Colbert, 1945) (Wilfarth, 1947). These ideas fall flat, however, due to the simple fact that there is no opening at the tip of the crest to let in air, and would not have been large enough to function as a sort of air reservoir for a dinosaur the size of Parasaurolophus. Other suggested uses for the crest have included the storage of a salt gland (Maryanska & Osmolska, 1979), an olfactory organ for smelling (Ostrom, 1962), and for thermoregulation (Wheeler, 1978).
The two most popular and well-established theories for the evolution of hollow crests in hadrosaurs, we now know, are visual display and sound production (Weishampel, 1981) (Evans, 2006). The hollow, winding tubes in lambeosaurine crests seem to have been useful in producing loud, resonating calls that could have carried for some distance. Hadrosaurs had a well-developed sense of hearing, which went hand-in-hand with their ability to produce loud noises (Weishampel, 1981).
The size and shape of the crest are features that help palaeontologists distinguish between the different species of Parasaurolophus. The most famous one is probably Parasaurolophus walkeri, the Albertan species. Ironically, this species is actually quite rare in comparison to other Albertan hadrosaurs, with only the one mostly complete holotype specimen and a few other fragmentary examples found so far (Evans et al., 2009). There’s also Parasaurolophus tubicen, a larger species from the Kirtland Formation of new Mexico with a deeper, straighter crest bearing more complicated passages within (Wiman, 1931) (Sullivan & Williamson, 1999).
A third species, Parasaurolophus cyrtocristatus*, comes from the younger Fruitland Formation of New Mexico and Kaiparowits Formation of Utah, and has a much shorter, more dramatically curled crest (Ostrom 1961). These crest shape differences have led to some palaeontologists suggesting that the type specimens of these different Parasaurolophus species represent either male of female (Weishampel, 1981). The idea is that the P. walkeri and P. tubicen type specimens, with their large and elaborate crests, were males, while the smaller-crested P. cyrtocristatus holotype was a female. It’s even been suggested that all these Parasaurolophus types are actually all the same species, and any difference between them is purely due to sexual dimorphism or growth stages (Paul, 2016). The idea that male hadrosaurs had larger, more visually impressive crests (presumably with greater resonating capabilities) than females of the same species is nothing new in dinosaur palaeontology, but is hard to prove, especially with small sample sizes of extinct animals. This idea doesn’t hold up any better with Parasaurolophus, since all three species are found in different locations and/or geologic formations.
*Update: shortly after this article was uploaded, a big new paper on Parasaurolphus cyrtocristatus was published (Gates et al., 2021). A couple new skulls of this species from the Fruitland and Kaiparowits Formations have surfaced are described in this new paper, and both preserve the back end of the skull much better than the fragmentary holotype. This preservation allows palaeontologists to study the proportions and arrangement of individual bones inthe skull of P. cyrtocristatus and therefore better define and diagnose the species without simple relying on crest shape and we’ve always been forced to do. Some interesting takeaways include the fact that the two American Parasaurolophus species, P. cyrtocristatus and P. tubicen are more closely related to each other than either is to the more northern P. walkeri. The authors could also now get a better understanding of how the bones in the skull form the unique crest of Parasaurolophus. They report that, despite the weird shape of the dinosaur’s headgear, the premaxilla bone which forms the tip of the upper jaws in most vertebrates dominates the top and sides of the crest in Parasaurolophus, a trait also seen in better known lambeosaurine genera like Corythosaurus.
There’s another hadrosaur that some have argued is a fourth species of Parasaurolophus. The enormous Charonosaurus jiayinensis from China had a similar, yet incomplete, skull to Parasaurolophus. An extensive 2014 paper found that Charonosaurus is actually so closely related to Parasaurolohpus that it should be considered the fourth species of that genus (Xing et al., 2014). Even though what we call Charonosaurus might not seem like such a big deal, since just about everyone agrees that it’s closely related to Parasaurolophus, this slight change in categorization comes with some interesting implications. Charonosaurus is far larger than any known Parasaurolophus, comes from a different continent, and dates back to the very end of the Cretaceous period, a time when lambeosaurines are thought to have gone extinct in North America. This would be a strange record of Parasaurolophus having migrated into Asia, attainting a much larger size, and surviving until the very end of the age of dinosaurs.
Despite the rarity of Parasaurolophus, we’ve still been able to learn a fair bit about this dinosaur thanks to some precious specimens and clever science. One thing that interests palaeontologists is how quickly dinosaurs grew, when features like crests and horns appeared in growing dinosaurs, and how they changed shape as they developed. The time in an animal’s life that certain features of anatomy appear help to indicate what their use might be, and the growth rate of dinosaurs allows us to figure out what their metabolisms were like. We have some understanding of this in Parasaurolophus thanks to a wonderfully complete juvenile specimen collected from Utah (Farke et al., 2013). This young specimen was just over 2 meters long when it died, only about a quarter the size it would likely reach as an adult. What’s interesting, though, is that it had already started to develop that characteristic crest. In this juvenile, however, the partially grown crest was low and round, and would presumably later elongate and curve backwards.
Let’s return to the original Parasaurolophus specimen though, the Royal Ontario Museum P. walkeri holotype. Some new (at the time of this writing) research has finally answered some old questions about this specimen, and revealed even more information about Parasaurolophus in general. The P. walkeri holotype is weird for a number of reasons. If you take quick glance at the vertebrae of the dinosaur, you’ll notice that the neural spines which stick out off of dorsal vertebrae (as in, the backbones between the neck and hips) 7 and 8 are weirdly bent away from each other. These deformed bones create a notch-like effect in the dinosaur’s back. In addition to that, an odd disc-shaped growth of bone is present on the top of the spine of dorsal vertebra 6. The tip of dorsal 7’s spine, being bent forward towards the back of dorsal 6, is fused to this bony disc. The dinosaur also has several healed injuries to its left ribs (the only well-preserved side of the animal), and a strange bony grown on the bottom of its ilium, one of the hip bones.
Injuries and infections in dinosaur bones are certainly not unheard of, and for some reason they seem to be especially common in hadrosaurs. But the Parasaurolophus walkeri holotype clearly has some weird extra stuff going on, pathology-wise. In a new paper (Bertozzo et al., 2020), a team of palaeontologists closely examined the battered old skeleton of the Toronto Parasaurolophus to figure out what the heck happened to this animal.
The suggestion they presented is as vivid as it is fun, and explains a lot of what we see in the dinosaur. The idea is that this Parasaurolophus, while it was still alive, was struck violently by something big and heavy. Maybe it was a larger dinosaur, or a falling rock, or maybe (as is wonderfully depicted in the paper), it was hit by a falling tree truck. It’s not an outlandish idea, and is something that happens to modern day animals from time to time. Perhaps, during a heavy tropical storm rolling in from the Western Interior Seaway, hurricane winds swept across the coastal plains, uprooting a large tree which came crashing down on the left side of a Parasaurolophus trying to find shelter in the squall.
It’s pretty much impossible to prove that this exact thing happened to this exact dinosaur, but it sparks the imagination, and the simplest explanation is indeed that something big did strike the animal on its top and left side, cracking its ribs in the process. Furthermore, the object striking the top-left of the dinosaur led to the strange growths on its vertebrae, causing the v-shaped notch and round bony growth. How did this happen? Well, let’s go back to Parks’ 1922 description of the specimen.
Parks’ idea of a fleshy frill present along the neck of Parasaurolophus partly came from the idea that a ligament, specifically the nuchal ligament which spans along the top of some animals necks, ran from the tip of the crest and connected to that strange disc-shaped growth on top of dorsal vertebra 6. This idea doesn’t actually work, since it would drastically restrict the movement of the dinosaur’s head, and in modern animals with this ligament it attaches to the back of the skull, and the crest of Parasaurolophus actually grows out of the snout and nose area. Nevertheless, this idea did spawn an abundance of Parasaurolophus depictions over the decades with a soft, fleshy frill running from the underside of the crest to around the creature’s shoulders or beyond.
Bertozzo et al., 2020 postulate that dorsal 6 is indeed the attachment point of a nuchal ligament in Parasaurolophus, and that the growth on top of that vertebra is the result of extra bone development due to trauma inflicted at the site by whatever hit the dinosaur. Furthermore, the stress on the nuchal ligament may have caused it to tighten and pull the neural spines near the front of the back forward. Hadrosaurs also had a mesh of ossified tendons along their dorsal side from the mid back to the base of the tail in order to stiffen the torso and keep the hips and tail upright. In this Parasaurolophus the ossified tendons may have yanked the neural spines of dorsal 7 and beyond backwards. All this yanking of vertebrae seems to have caused that conspicuous notch to form. The overgrowth of bone in the hip, Bertozzo et al. suggest, could be the result of the dinosaur walking with a limp after the initial injury. Indeed, because of the fact that the rib fractures were healed, we know the Parasaurolophus survived for at least months, if not years, after it was inured, likely in considerable pain and discomfort.
So that’s the life of one Parasaurolophus. What about this type of dinosaur in general- what else does science know about Parasaurolophus? Due to its relative rarity, we don’t know quite as much about Parasaurolophus as we do more common hadrosaurs like Corythosaurus or Edmontosaurus, but there’s still some interesting bits of info out there, such as the life appearance of the animal. Bertozzo et al. didn’t just interpret the injuries to the P. walkeri holotype, they also figured out, roughly, how deep the dinosaur’s neck was in life. Having figured out which vertebra the nuchal ligament attached to, they reasoned that it connected to the back of the skull as it does in living reptiles. This told them more or less where the dorsal margin of the neck would’ve been. As is suggested by ‘mummified’ duckbills that preserve skin impressions from this region, we can now safely say that most if not all hadrosaurs had much deeper, well-muscled necks than what’s suggested by the vertebrae of that region alone.
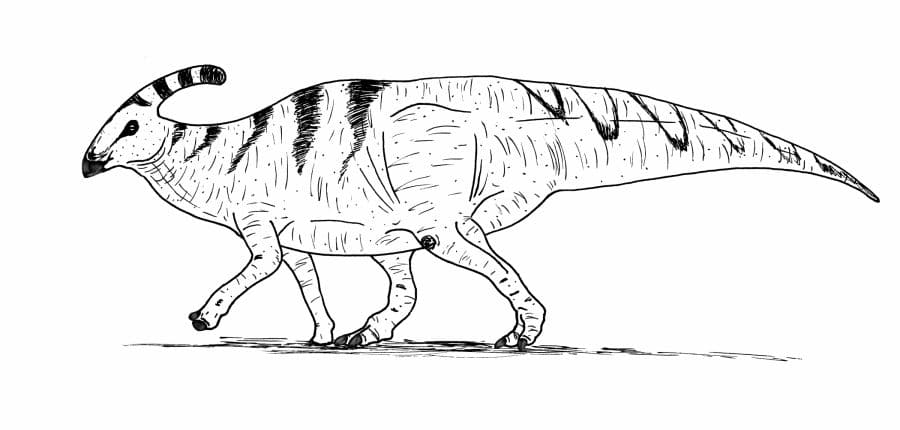
A somewhat conservative reconstruction of Parasaurolophus walkeri, following Bertozzo et al., 2020. By Nicholas Carter, based on the skeletal diagram by Scott Hartman.
Speaking of skin impressions, in hadrosaurs they aren’t uncommon, and certain specimens of different species have even been found with amazingly extensive patches surrounding the bones. In his 1922 description, Parks noted the presence of a small patch of skin impressions found with Parasaurolophus walkeri, which was he wrote contained fairly small, non-overlapping scales, but with no trace of the interspersed larger tubercles found in other duckbills.
Like all other known hadrosaurs, Parasaurolophus was a generalist herbivore, and likely took advantage of whatever edible vegetation struck its fancy. Far from the grazer of soft aquatic vegetation that palaeontologists of past generations imagined it to be, Parasaurolophus and its other hadrosaur relatives were mostly land-based and fed on anything from ferns to conifer twigs to even tree bark, thanks to their cropping beaks and grinding teeth (Chin, 2007). The tall build, and ability to rear up, allowed hadrosaurs to access tree branches and other such sources of food that were out of reach of the ceratopsids, ankylosaurs, and smaller herbivores they shared their habitat with (Mallon et al., 2013). This ability to exploit food resources from up in trees would’ve been handy in an environment devoid of sauropods such as the Dinosaur Park Formation.
All three North American species of Parasaurolophus lived in the warm, lush flats between the Rocky Mountains and inland seaway that existed during the late Cretaceous. Certain species at particular times could be found in southern Alberta, Utah, and New Mexico. The lower Dinosaur Park Formation, where P. walkeri is found, was at the time a swampy coastal region with wooded lowlands and floodplains cut by winding rivers. It’s been informally suggested that perhaps Parasaurolophus is uncommon here because it may have preferred drier, upland environments to the west, and spent less time in the coastal bayous than other hadrosaurs. That said, other species of this genus are more common in somewhat similar habitats to the south, so perhaps the Albertan Parasaurolophus specimens represent individuals near the limit of their northern range.
The social behavior of Parasaurolophus is another mystery, despite it being depicted as a herding animal in the Jurassic Park films. Bonebeds that likely represent herds have been found from both lambeosaurine and saurolophine dinosaurs, but these mass accumulations are unknown in Parasaurolophus so far. That said, it does seem unlikely that a genus would evolve a structure like the visually and audibly striking crest of Parasaurolophus if it didn’t spend at least some of its time in groups.
Who knows what else we’ll discover about Parasaurolophus in the years to come. We can only hope that more well-preserved fossils of this popular yet rare genus are found in the Cretaceous rocks of western North America. It fills the mind with wonder to look out onto the vast prairie badlands and think that this strange and unique creature called this place home, millions of years ago.

References
Bertozzo, F., Manucci, F., Dempsey, M, Tanke, D H., et al. Description and etiology of paleopathological lesions in the type specimen of Parasaurolophus walkeri (Dinosauria: Hadrosauridae), with proposed reconstructions of the nuchal ligament J. Anat. 2020; 00: 1– 15. https://doi.org/10.1111/joa.13363
Chin, K. (September 2007). “The Paleobiological Implications of Herbivorous Dinosaur Coprolites from the Upper Cretaceous Two Medicine Formation of Montana: Why Eat Wood?”. PALAIOS. 22 (5): 554. Bibcode:2007Palai..22..554C. doi:10.2110/palo.2006.p06-087r. S2CID 86197149
Colbert, Edwin H. (1945). The Dinosaur Book: The Ruling Reptiles and their Relatives. New York: American Museum of Natural History, Man and Nature Publications, 14. p. 156.
Evans, D.C. (2006). “Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs”. Paleobiology. 32 (1): 109–125.
Evans, D.C.; Bavington, R.; Campione, N.E. (2009). “An unusual hadrosaurid braincase from the Dinosaur Park Formation and the biostratigraphy of Parasaurolophus (Ornithischia: Lambeosaurinae) from southern Alberta”. Canadian Journal of Earth Sciences. 46 (11): 791–800.
Farke, A.A.; Chok, D.J.; Herrero, A.; Scolieri, B.; Werning, S. (2013). Hutchinson, John (ed.). “Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids”. PeerJ. 1: e182. doi:10.7717/peerj.182. PMC 3807589. PMID 24167777.
2021. Description and rediagnosis of the crested hadrosaurid (Ornithopoda) dinosaur Parasaurolophus cyrtocristatus on the basis of new cranial remains. PeerJ 9:e10669 https://doi.org/10.7717/peerj.10669
Parks, W.A. (1922). “Parasaurolophus walkeri, a new genus and species of trachodont dinosaur”. University of Toronto Studies: Geological Series. 13: 5–32.
Mallon, Jordan C.; Evans, David C.; Ryan, Michael J.; Anderson, Jason S. (2013). “Feeding height stratification among the herbivorous dinosaurs from the Dinosaur Park Formation (upper Campanian) of Alberta, Canada”. BMC Ecology. 13: 14. doi:10.1186/1472-6785-13-14. PMC 3637170. PMID 23557203.
Maryanska, T.; Osmólska, H. (1979). “Aspects of hadrosaurian cranial anatomy”. Lethaia. 12 (3): 265–273.
Ostrom, J.H. (1961). “A New Species of Hadrosaurian Dinosaur from the Cretaceous of New Mexico”. Journal of Paleontology. 35 (3): 575–577.
Ostrom, John H. (1962). “The cranial crests of hadrosaurian dinosaurs”. Postilla. 62: 1–29.
Paul, Gregory S. (2016). The Princeton Field Guide to Dinosaurs 2nd Edition. United States of America: Princeton University Press. pp. 341-342
Romer, Alfred Sherwood (1933). Vertebrate Paleontology. University of Chicago Press. p. 491
Sternberg, Charles M. (1935). “Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous”. Canada Department of Mines Bulletin (Geological Series). 77 (52): 1–37.
Sullivan, R.S.; Williamson, T.E. (1999). “A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus” (PDF). New Mexico Museum of Natural History and Science Bulletin. 15: 1–52.
Weishampel, D.B. (1981). “Acoustic Analysis of Vocalization of Lambeosaurine Dinosaurs (Reptilia: Ornithischia)” (PDF). Paleobiology. 7 (2): 252–261.
Wheeler, P.E. (1978). “Elaborate CNS cooling structure in large dinosaurs”. Nature. 275 (5679): 441–443.
Wilfarth, Martin (1947). “Russeltragende Dinosaurier”. Orion (Munich) (in German). 2: 525–532.
Wiman, C. (1931). “Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico”. Nova Acta Regia Societas Scientarum Upsaliensis, Series 4 (in German). 7 (5): 1–11.
Xing, H.; Wang, D.; Han, F.; Sullivan, C.; Ma, Q.; He, Y.; Hone, D.W.E.; Yan, R.; Du, F.; Xu, X. (2014). Evans, David C. (ed.). “New Basal Hadrosauroid Dinosaur (Dinosauria: Ornithopoda) with Transitional Features from the Late Cretaceous of Henan Province, China”. PLOS ONE. 9 (6): e98821. Bibcode:2014PLoSO…998821X. doi:10.1371/journal.pone.0098821. PMC 4047018. PMID 24901454.