
Huge dinosaurs like Pachyrhinosaurus and Edmontosaurus roamed the Grande Prairie area about 70 million years ago, but such heavyweights never had the Cretaceous world to themselves. There were plenty of smaller dinosaurs around, like the little carnivore Boreonykus, and dinosaurs were only one component of a diverse ecosystem that also included fish, turtles, mammals and many other vertebrates, alongside plants and invertebrates. However, small vertebrates can be frustratingly elusive in the fossil record, precisely because of their pint-sized proportions. Their delicate skeletons are easily destroyed before they can be fossilised – if not by a predator that gulps them down bones and all, then by scavengers and the elements. Comparatively few ever make it into the hands of palaeontologists.
In a scientific paper that came out just last November, a team of authors led by my former student Sam Hamilton described a skull bone that provides important information about one relatively small vertebrate that coexisted with the likes of Pachyrhinosaurus, namely a lizard. The specimen was collected from a site called the DC Bonebed on the north bank of the Wapiti River, by a field team from the University of Alberta, the Philip J. Currie Dinosaur Museum, the University of New England in Australia, and the University of Bologna in Italy, operating with valuable logistical support from Northwestern Polytechnic (then Grande Prairie Regional College).

Sam Hamilton, lead author of the fossil lizard paper, excavates the hard way at the DC Bonebed in 2018. Photo by Corwin Sullivan.
The DC Bonebed has yielded a mixed bag of individual bones and teeth from small to medium-sized vertebrates, but only two of them demonstrably belong to lizards, the other being an ankle bone called the astragalocalcaneum. The CBC’s Dennis Kovtun published a nice article about our research on these specimens, but I thought I’d take the opportunity to give a fuller account here.

Lizard frontal from the DC Bonebed, in top view (left) and side view (right). Scale bar: 1 cm. Photos by Robin Sissons.

Lizard astragocalcaneum from the DC Bonebed. Scale bar: 1 cm. Photo by Sam Hamilton.
Both the skull bone and the astragalocalcaneum are from the right side of the body, and are large by lizard standards. They were found on two successive days in the summer of 2017, as I realised only after a question from Dennis about when they had been discovered sent me to my field records, so chances are that they were very close together in the bonebed. Even so, there’s no guarantee that they represent the same individual, or even species.
Lizard astragalocalcanea are difficult to identify in isolation, so my coauthors and I couldn’t do much with that specimen beyond just reporting its existence, but the skull piece was another matter. It soon dawned on us that we had a bone called the frontal, which makes up the forehead in humans and the central part of the top side of the skull in reptiles.
The most important clue to the identification of the specimen is the presence of small pieces of bone, called osteoderms, on the upper surface of the frontal. Osteoderms, which form in the skin and act as defensive armour, are not uncommon in lizards but differ in appearance among lizard groups. The ones attached to the DC frontal are large, roughly polygon-shaped, and covered in pits separated by thin ridges. A high-resolution CT scan showed that they were fused to the underlying frontal, not just stuck to it.
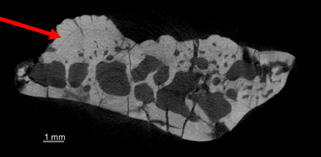
Cross-sectional image of lizard frontal, produced from CT scan data. Red arrow indicates an osteoderm. Note the lack of separation between the osteoderm and the frontal itself.
Osteoderms of the type seen in the DC frontal are characteristic of one particular lizard group, which (believe it or not!) is formally known to science as the Monstersauria. Monstersaurs come by their whimsical name honestly, in that the most (in)famous living member of the group is Heloderma suspectum, known in English as the Gila monster. Other living monstersaurs are also classified within the genus Heloderma, and are called beaded lizards. This term indirectly refers to the osteoderms, which give the skin of all Heloderma species a knobbly appearance.

A Gila monster draped over a rock in its enclosure at the Bronx Zoo in New York, USA. Photo by Corwin Sullivan.
A couple of other features of the frontal corroborate the interpretation that it belongs to a monstersaur. In many lizard groups the right and left frontals fuse together, but our right frontal was clearly separate from its left side counterpart, as in monstersaurs. Furthermore, the wall of bone attached to the underside of the frontal, called the subolfactory process, was well developed but not so large that it would have contacted the subolfactory process of the opposite frontal. This type of mid-sized subolfactory process is far from universal among lizards, but quite usual in Cretaceous monstersaurs.

Lizard frontal from the DC Bonebed, in bottom view. Scale bar: 1 cm. Red arrow indicates the subolfactory process. Note that the subolfactory process would not have extended to the skull midline (indicated by red dashes) and therefore would not have contacted the subolfactory process of the opposite frontal. Photo by Robin Sissons.
The frontal of what my coauthors and I affectionately call the DC Monster bears an especially close resemblance to the same bone in a partial monstersaur skeleton from the Cretaceous of Montana, which was described in an unpublished thesis by Michael Balsai back in 2001 but has never been formally reported in the scientific literature. The frontals of the Montana specimen are very similar to the DC frontal, even down to some details of the osteoderm arrangement.
By contrast, a partial frontal that was collected from White Rock Coulee in the 1970s, and identified in a 1996 paper by Keqin Gao and Dick Fox as belonging to a species called Labrodioctes montanensis, looks quite different. The osteoderms on the Labrodioctes frontal have straighter edges and sharper corners than those on the DC and Montana frontals, and are also separated by better-defined grooves and only patchily covered in the typical monstersaurian ridge-and-pit pattern, with some areas showing only scattered pitting.

Lizard frontal from White Rock Coulee, identified as Labrodioctes montanensis, in top view (left) and side view (right). Scale bar: 1 cm. Photos by Robin Sissons.
Michael’s thesis boldly declared the Montana specimen to belong to Palaeosaniwa canadensis, a monstersaur species named in 1928 by the American palaeontologist Charles Whitney Gilmore. Many individual bones from the Late Cretaceous of North America have been identified as Palaeosaniwa over the years, but it’s fair to say that Palaeosaniwa is rather ill-defined. The only characteristic that may reliably distinguish Palaeosaniwa from other monstersaurs is a groove on the underside of the trunk vertebrae, but whether the groove is really restricted to Palaeosaniwa or is actually more widespread is uncertain. The trunk vertebrae of the Montana specimen show the ventral groove, but my coauthors and I consider this to be only limited evidence for identifying the specimen as Palaeosaniwa canadensis.
The importance of taxonomic names shouldn’t be exaggerated. What is clear is that the DC Monster likely represents the same species as the Montana specimen. Whether this species should be called Palaeosaniwa canadensis or something else is really just a taxonomic formality, but it’s definitely different from the species represented by the Labrodioctes frontal. Accordingly, there were at least two different monstersaurs in western North America during the part of the Late Cretaceous to which all three specimens can be dated, called the Campanian Age.
It’s possible to make some educated guesses about the size, habitat and lifestyle of the DC Monster. The DC frontal is about 35 mm long, and the frontals of the Montana specimen more like 40 mm. This compares to 39 mm for the frontals of a specimen of the Mongolian Cretaceous monstersaur Estesia mongoliensis, which had a total skull length of 150 mm. The DC Monster probably had a skull length in the same ballpark, as do young adults of the modern Komodo dragon. The young Komodos have total body lengths of about 1.5 metres, so perhaps the DC Monster was about that size as well, though such estimates are very inexact.
In the modern world, the only reptile found in the Grande Prairie area is the red-sided garter snake Thamnophis sirtalis, and big lizards are confined to much lower latitudes. The Gila monster occurs no farther north than southern Nevada and Utah, at a latitude of about 38° N. Grande Prairie is at 55° N, and during the Campanian was probably just outside the Arctic Circle, at more like 63° N. The subarctic might seem like a strange place for a lizard the size of the DC Monster, but the Campanian climate was much warmer than today’s, so that large cold-blooded reptiles were able to thrive even at high latitudes. Nevertheless, what’s now Grande Prairie would still have had short days in winter and long days in summer, so the environment would have been strongly seasonal. This dovetails interestingly with the tendency of the modern Heloderma species to favour seasonal environments, including the seasonally dry tropical forest of western Mexico and the deserts and scrublands of the southwestern United States.
Like other living and fossil monstersaurs, the Montana specimen has sharp teeth, implying that the DC Monster was a carnivore. The Gila monster and beaded lizards are notorious for having glands in their lower jaws that produce strong venom, which flows into the mouth via grooves in the lower teeth. These lizards can therefore defend themselves with a toxic bite, which is rarely if ever fatal to humans but can be horribly painful and cause faintness, a drop in blood pressure, and other serious symptoms. Estesia has venom grooves as well, but the lower teeth of the Montana specimen appear to lack them, which suggests that the DC Monster would have had to rely on osteoderms and a good strong bite for protection in the dangerous world of the Campanian.
The frontal from the DC Bonebed isn’t quite the first evidence that monstersaurs inhabited the Grande Prairie area in Campanian time. In a richly descriptive 2008 paper on Pachyrhinosaurus lakustai from the Pipestone Creek Bonebed, Phil Currie and his coauthors included a Palaeosaniwa-like vertebra in a list of vertebrate fossils not belonging to Pachyrhinosaurus that had been recovered from the site. However, the DC frontal provides decisive new evidence for the presence of monstersaurs, and establishes a strong, specific link to the Montana specimen while conversely showing important differences from the Labrodioctes frontal from White Rock Coulee. Not bad for one skull bone from a little monster!